NRF2和AKT激活或FBP1缺失在肝細胞衰老和肝癌進展中的作用_abio生物試劑品牌網
喜報!恭喜我們的客戶使用 MCE 產品在 Nature 發表創新性研究~
本文要和大家分享的是近期發表在 Nature 的客戶文獻: “FBP1 controls liver cancer evolution from senescent MASH hepatocytes” (FBP1 控制衰老的 MASH 肝細胞的肝癌進化)[1]。
該文獻使用了 Etoposide (HY-13629)、5-azacytidine (HY-10586)、BpV (HOpic) (HY-128693)、NK252 (HY-19734) 進行研究,其中 Etoposide、5-azacytidine 常用于誘導細胞自噬,BpV 是 PTEN 抑制劑,NK-252 是一種有效的的 Nrf2 激活劑。

該研究揭示了 FBP1 在肝癌 (HCC) 發生中的作用,發現其在衰老樣 MASH 肝細胞中升高,MASH 中 FBP1 和 p53 抑制肝癌進展,后兩者下調則促進。FBP1 缺失通過 AKT 激活促進肝癌發生,NRF2 與 FBP1 相互作用影響肝癌相關過程,包括代謝和細胞周期基因表達等。
Section.01
背景介紹
Nature 子刊 Gastroenterology & Hepatology 發表的一篇綜述指出:非酒精性脂肪性肝病 (NAFLD) 全球患病率為 20%~25%。而我國上海、北京等地區的流行病學調查顯示,10 年間,普通成人 B 超診斷的非酒精性脂肪肝患病率從 15% 增加到 31% 以上,也就是說,每三個人中就有一個是脂肪肝。NAFLD 影響了全球多達三分之一的人口,且已成為肝硬化、肝衰竭和肝癌發病率快速增長的主要原因[1][2]。
代謝功能障礙相關脂肪性肝炎 (MASH) 是脂肪性肝病常見的炎癥并發癥,盡管 MASH 能夠觸發肝細胞衰老死亡,但這種狀態卻同時增加了 HCC 的風險。MASH 是如何在增加肝細胞癌風險的同時又抑制肝細胞癌的發生?
Section.02
研究方法與發現
人類 HCC 中的 p53 和 FBP1 表達下降
研究人員發現,FBP1 (果糖-1,6-二磷酸酶 1) 在調控肝細胞衰老和 HCC 發展中扮演著至關重要的角色。正常情況下,FBP1 有助于失活促癌酶 AKT,并維持 TP53 (一種重要的腫瘤抑制蛋白) 的水平。當 FBP1 表達減少時,AKT 被激活,加速了 TP53 的降解,并穩定了促癌轉錄因子 NRF2,從而促進了癌癥的發展 (圖 1)。
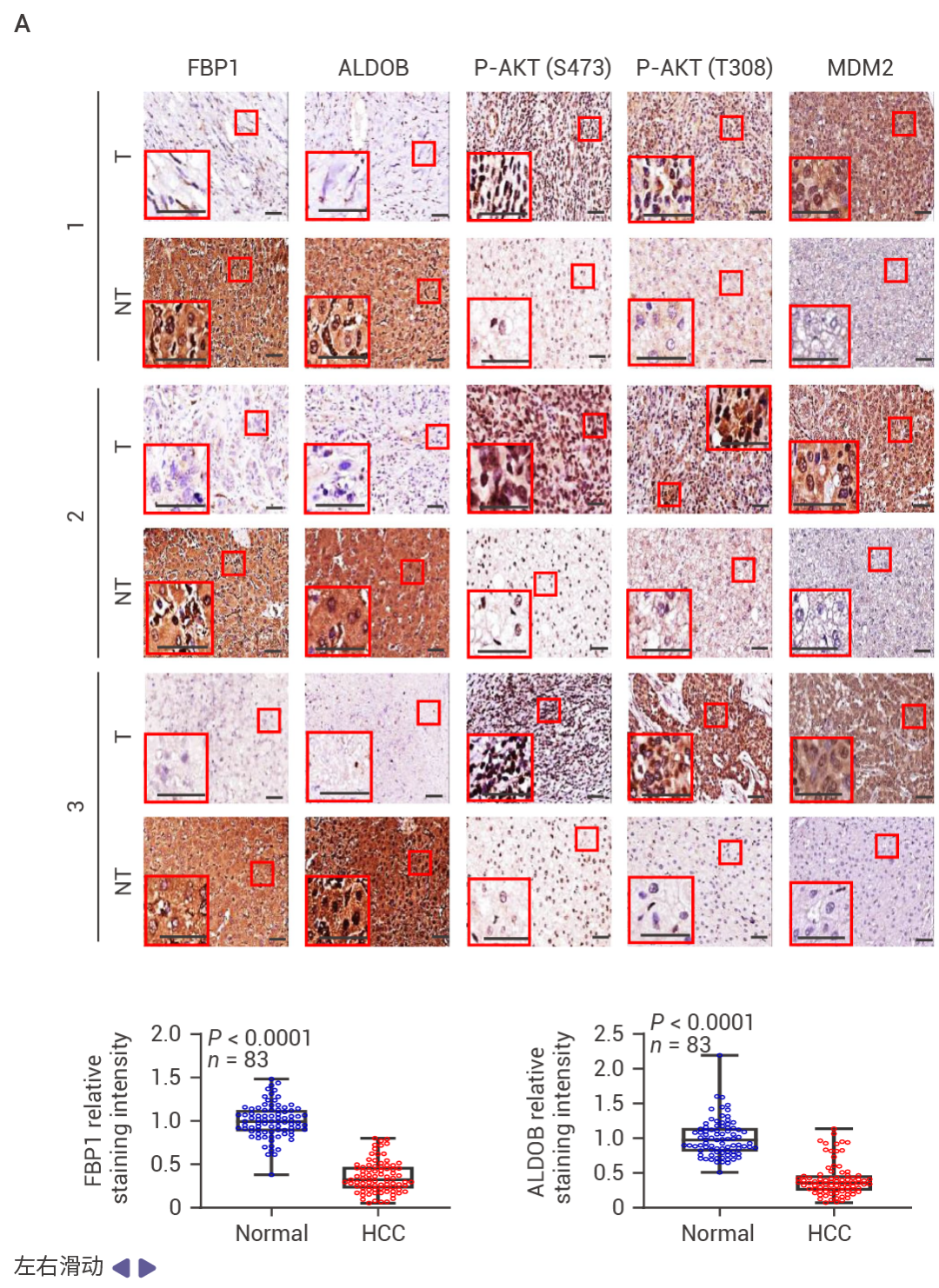
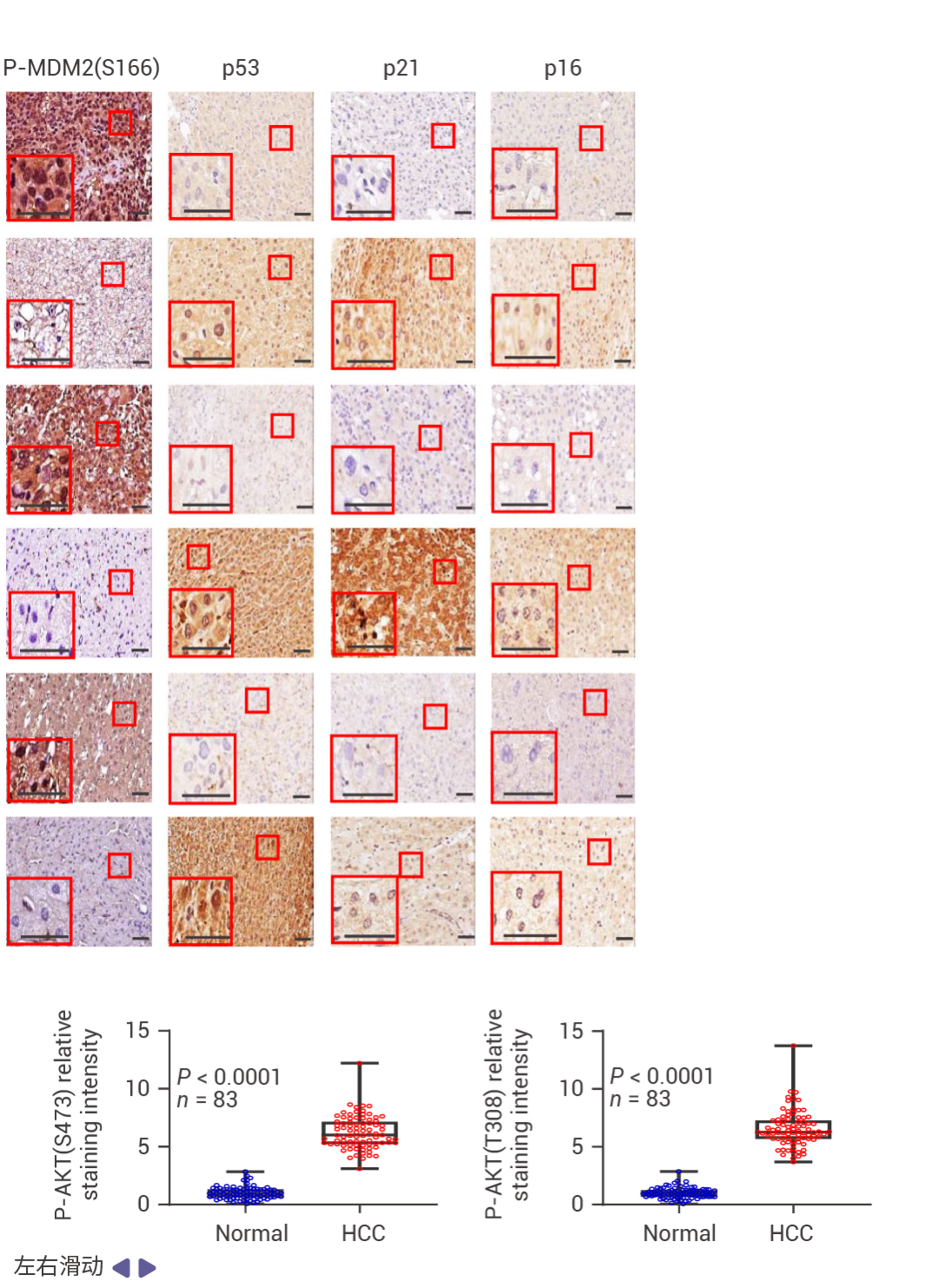
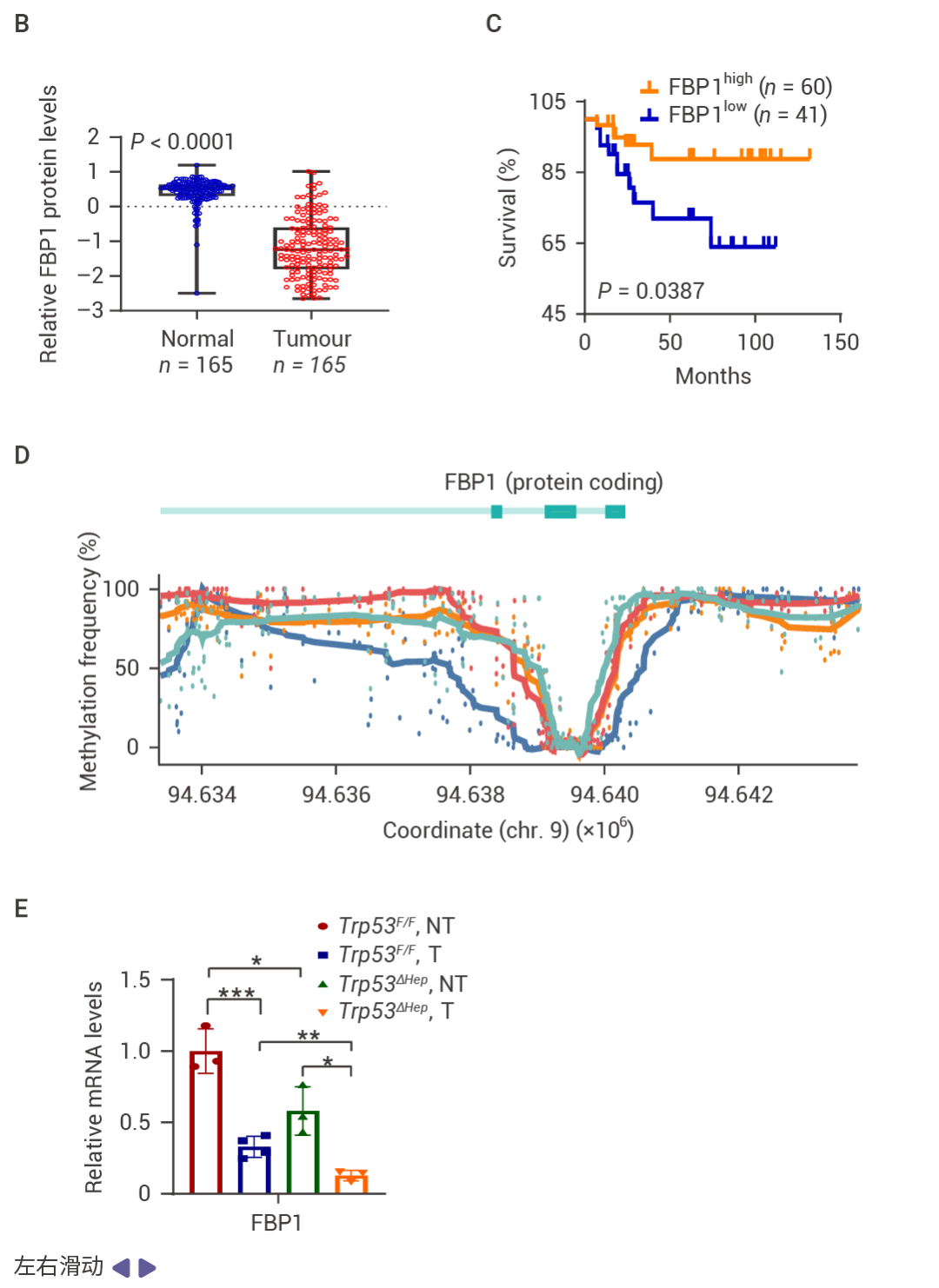 圖 1. FBP1 和 P53 在人類 HCC 中表達下調。
圖 1. FBP1 和 P53 在人類 HCC 中表達下調。
在 83 例 HCC 中,71% 的病例中 FBP1 表達下調,并且與 ALDOB 丟失、AKT 激活、MDM2 Ser166 (AKT 位點) 磷酸化升高以及 p53、p21CIP1 和 p16INK4a 表達下降顯著相關 (圖 1a)。在 HCC 中,FBP1 廣泛下調 (圖 1b)。低 FBP1 和 ALDOB 表達與較差的生存率相關 (圖 1c)。甲基化組分析顯示,與正常肝細胞核相比,HCC 細胞核中 FBP1 啟動子區域的高甲基化水平,而 TP53 啟動子的甲基化未見改變 (圖 1d)。TP53 缺失降低了小鼠肝臟、致癌物誘導的 HCC、人類肝細胞以及 TP53 野生型 HCC 細胞系 HepG2 和 SK-Hep-1 中 FBP1 的 mRNA 和蛋白水平 (圖 1e)。
MASH 提高的 FBP1 和 p53 抑制 HCC
盡管肝細胞衰老被認為是推動 MASH 和纖維化的因素,但敲除衰老細胞中的 FBP1 會增加肝纖維化、肝脂肪變性、肝臟和血脂,并抑制糖原積累和胰島素抵抗。肝細胞衰老先于 MASLD/MASH,但它抑制了纖維化和 HCC 的進展,Trp53ΔHep 小鼠脂肪肝炎的升高也表明了這一點。DNA 損傷誘導的 FBP1 和 p53 在 MASH 中顯著上調,提示其在 MASH 病理過程中可能發揮重要作用 (圖 2)。
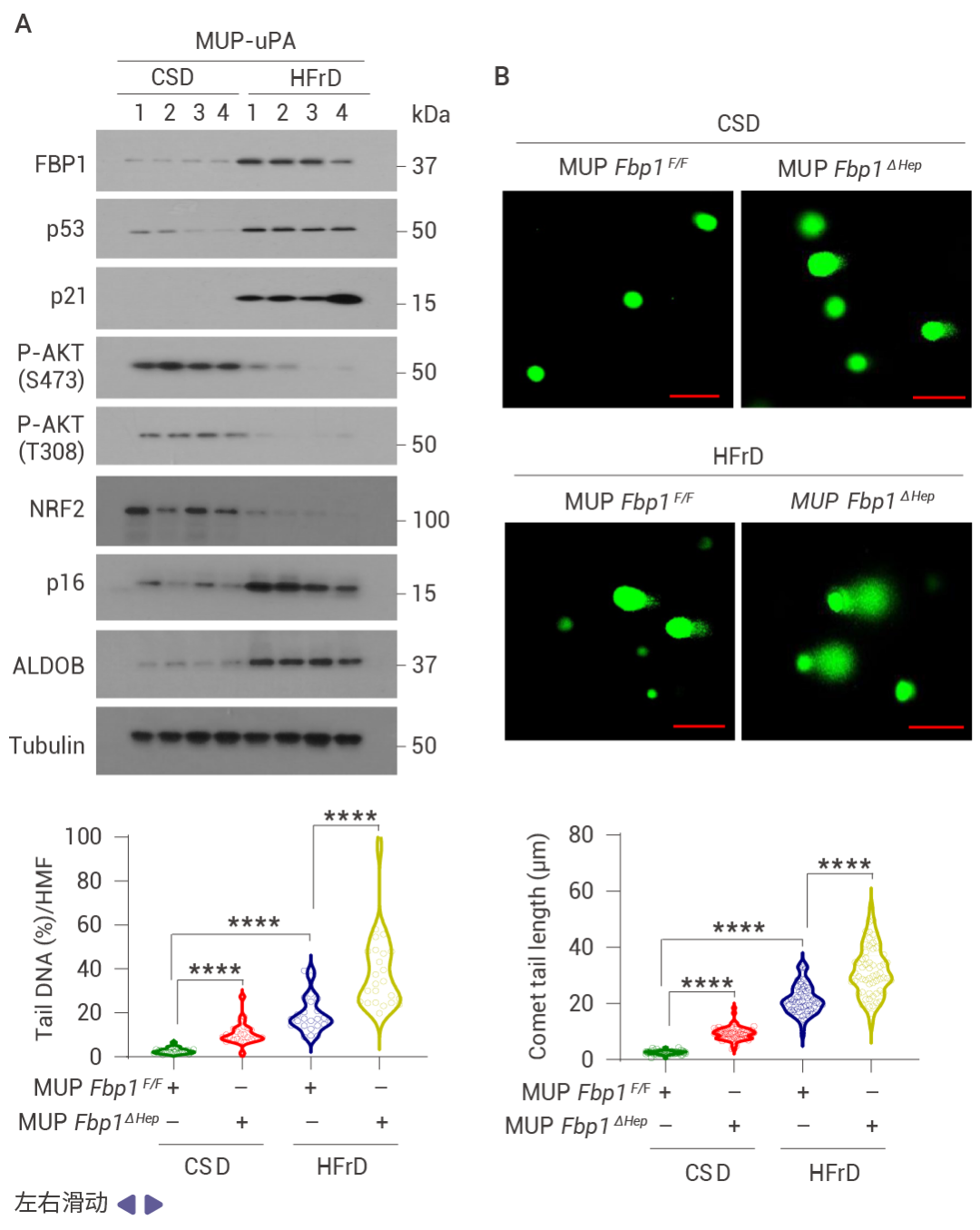
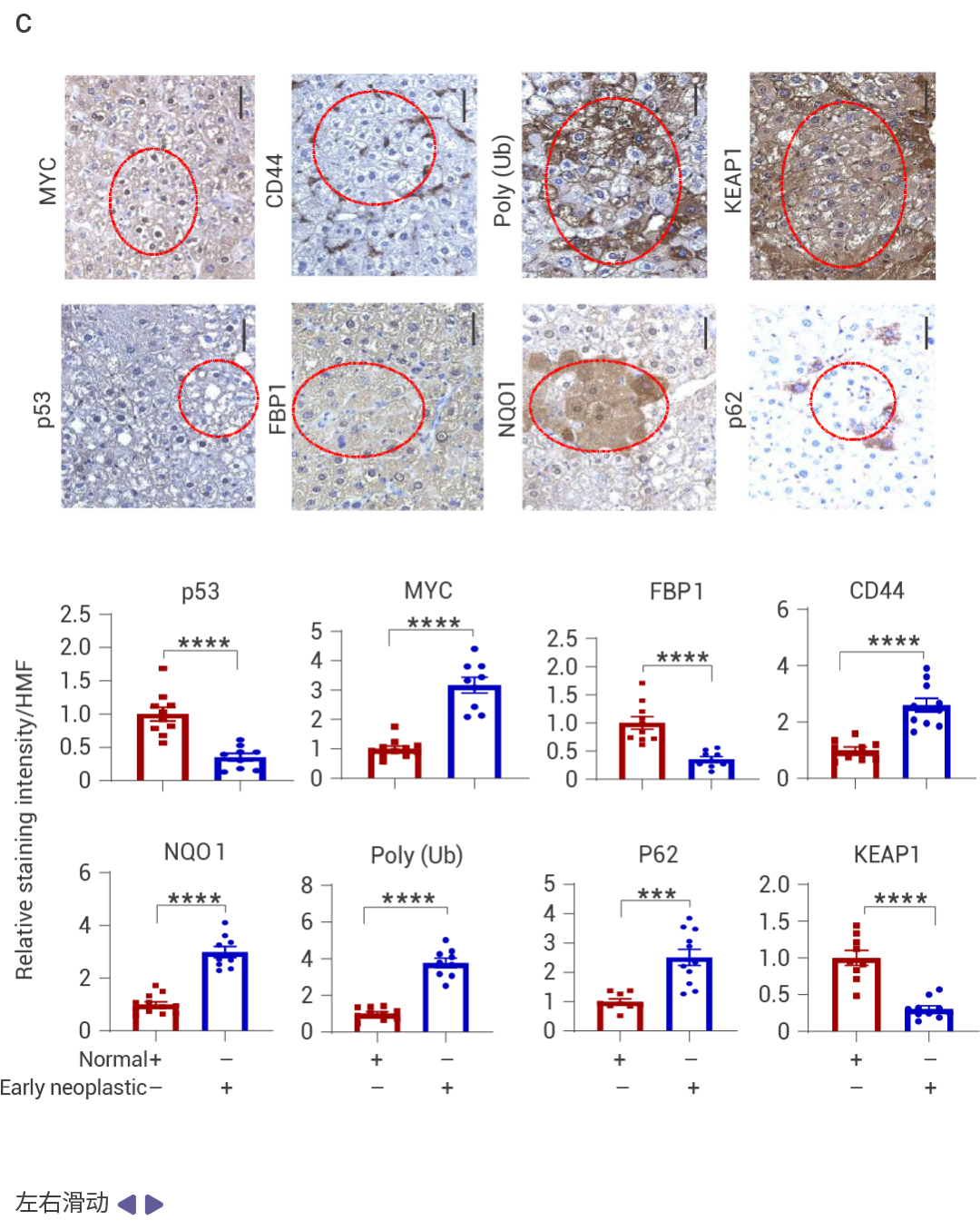
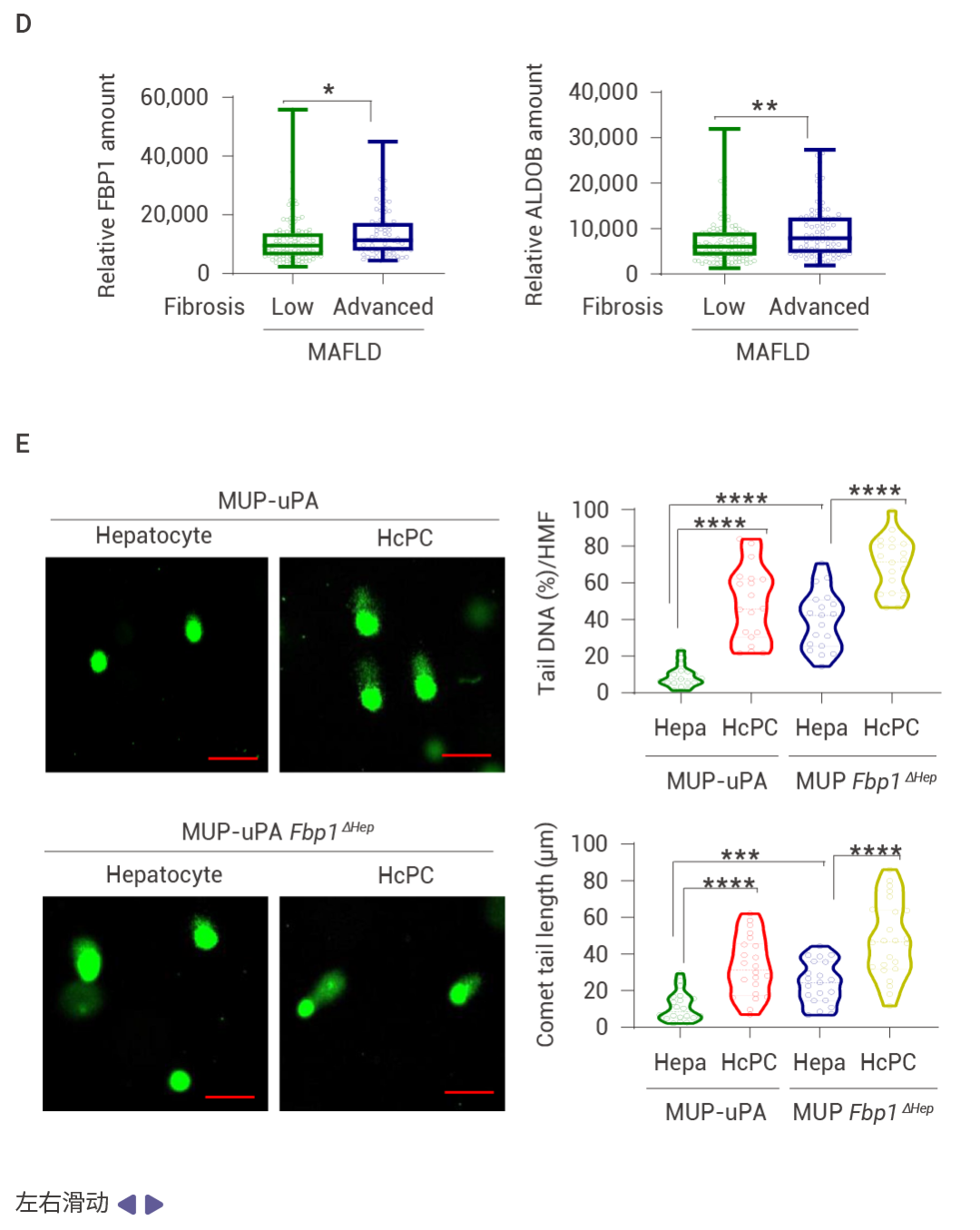
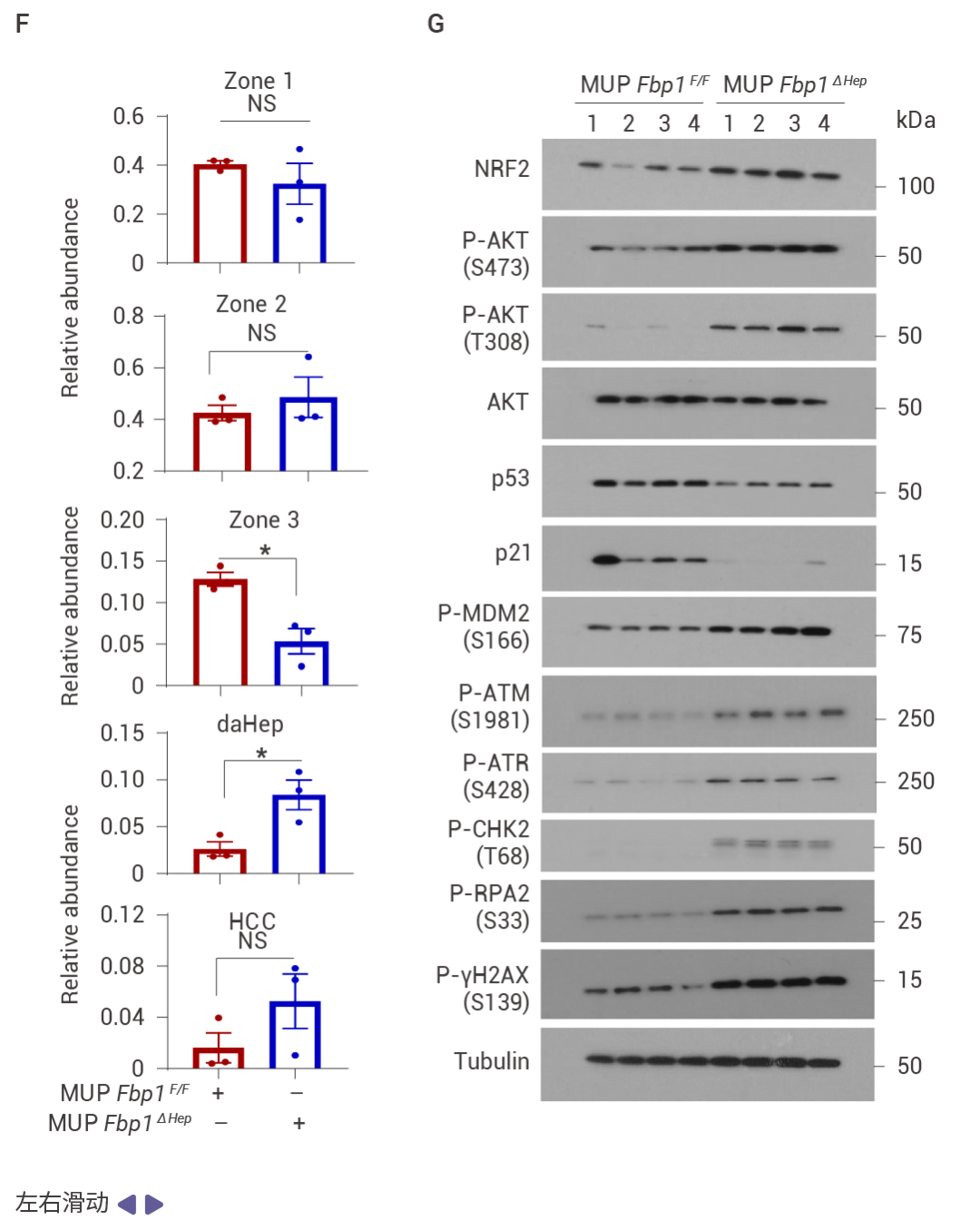 圖 2. DNA 損傷誘導的 FBP1 和 p53 在 MASH 中的上調。
圖 2. DNA 損傷誘導的 FBP1 和 p53 在 MASH 中的上調。
喂食易感 MASH 的 MUP-uPA 小鼠高果糖飲食 (HFrD),而非等熱量的玉米淀粉飲食 (CSD),誘導了 p53、p21CIP1、p16INK4a 和 FBP1 的表達,并抑制了 AKT 激活 (圖 2a)。肝細胞 DNA 損傷及其 DNA 損傷反應 (DDR) 介質升高,同時觀察到 P-RPA2 和衰老相關的 β-半乳糖苷酶表達升高 (圖 2b)。HFrD 飼喂誘導了早期癌前病變,這些病變中聚泛素化蛋白聚集體和自噬伴侶蛋白 p62 的積累提示自噬受損,NRF2 (NQO1)、CD44 和 MYC 被激活,而 p53 和 FBP1 表達下調 (圖 2c)。晚期人類 MASH 蛋白質組和組織的分析證實了 FBP1、ALDOB、p53、p21CIP1、p16INK4a 和 DDR 的上調,以及 AKT 和 NRF2 活性的降低 (圖 2d)。FBP1 的缺失增加了 HCPC 的數量,而 HcPC 比未聚集的肝細胞表現出更多的 DNA 損傷,同時伴隨著 DDR 和 DNMT1 的上調、AKT 和 NRF2 的激活,以及 FBP1、p53、p21CIP1 和 p16INK4a 的下調 (圖 2e)。單核 RNA 測序表明 FBP1 缺失增加了疾病相關肝細胞的數量,同時減少了 3 區肝細胞 (圖 2f)。WB 結果表明 FBP1 缺失緩解了 HFrD 誘導的 AKT 抑制,增強了 GSK3β Ser9 和 MDM2 Ser166 的磷酸化、DDR 標志物的表達,并下調了 p53 和 p21CIP1 (圖 2g)。
FBP1 的缺失通過 AKT 激活促進腫瘤發生
研究表明 FBP1 或 p53 的缺失以及 AKT 的激活均能促進 NRASG12V 驅動的腫瘤發生,而通過恢復 FBP1 功能或抑制 AKT 信號通路可以顯著抑制腫瘤的發生和發展 (圖 3)。
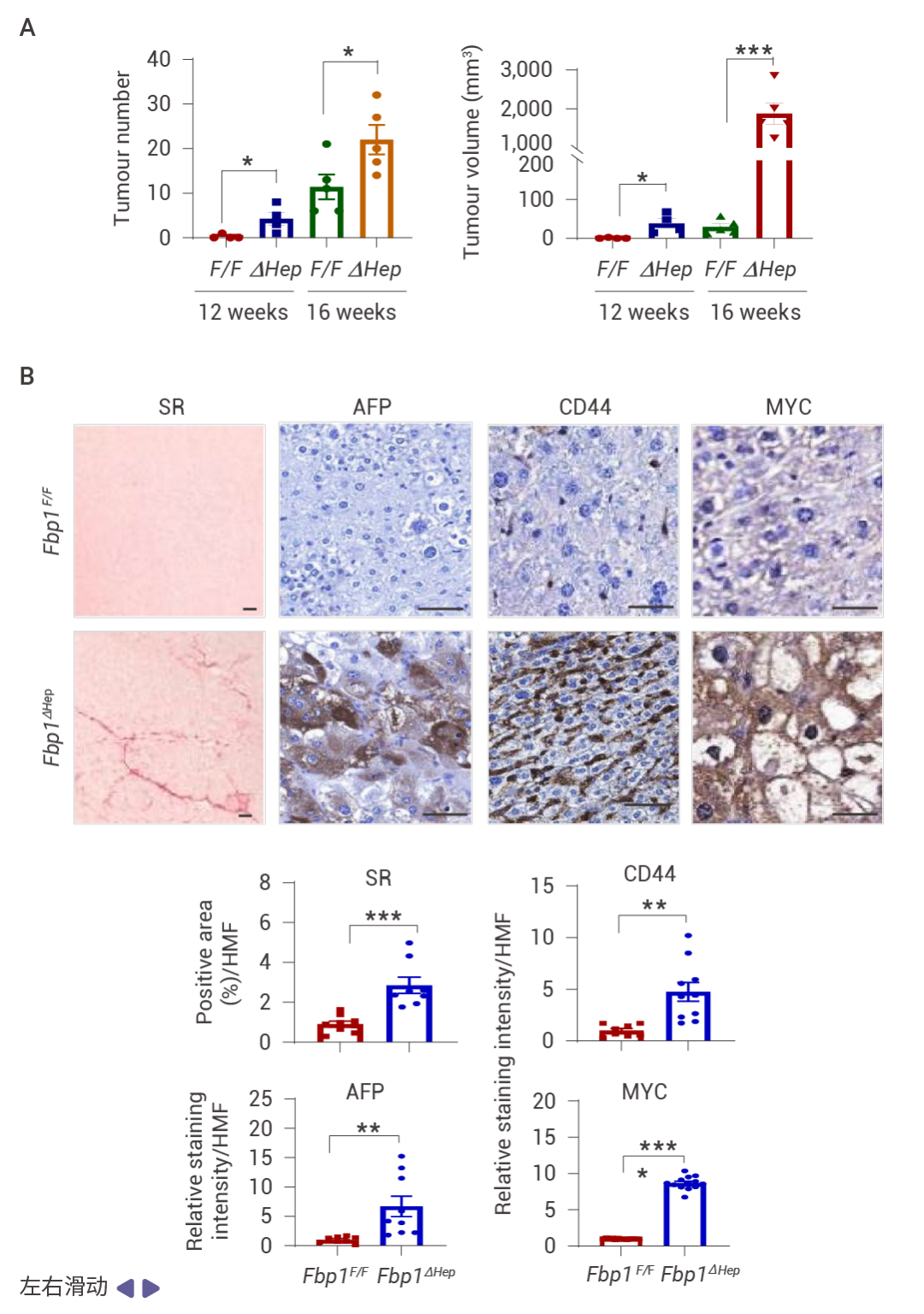
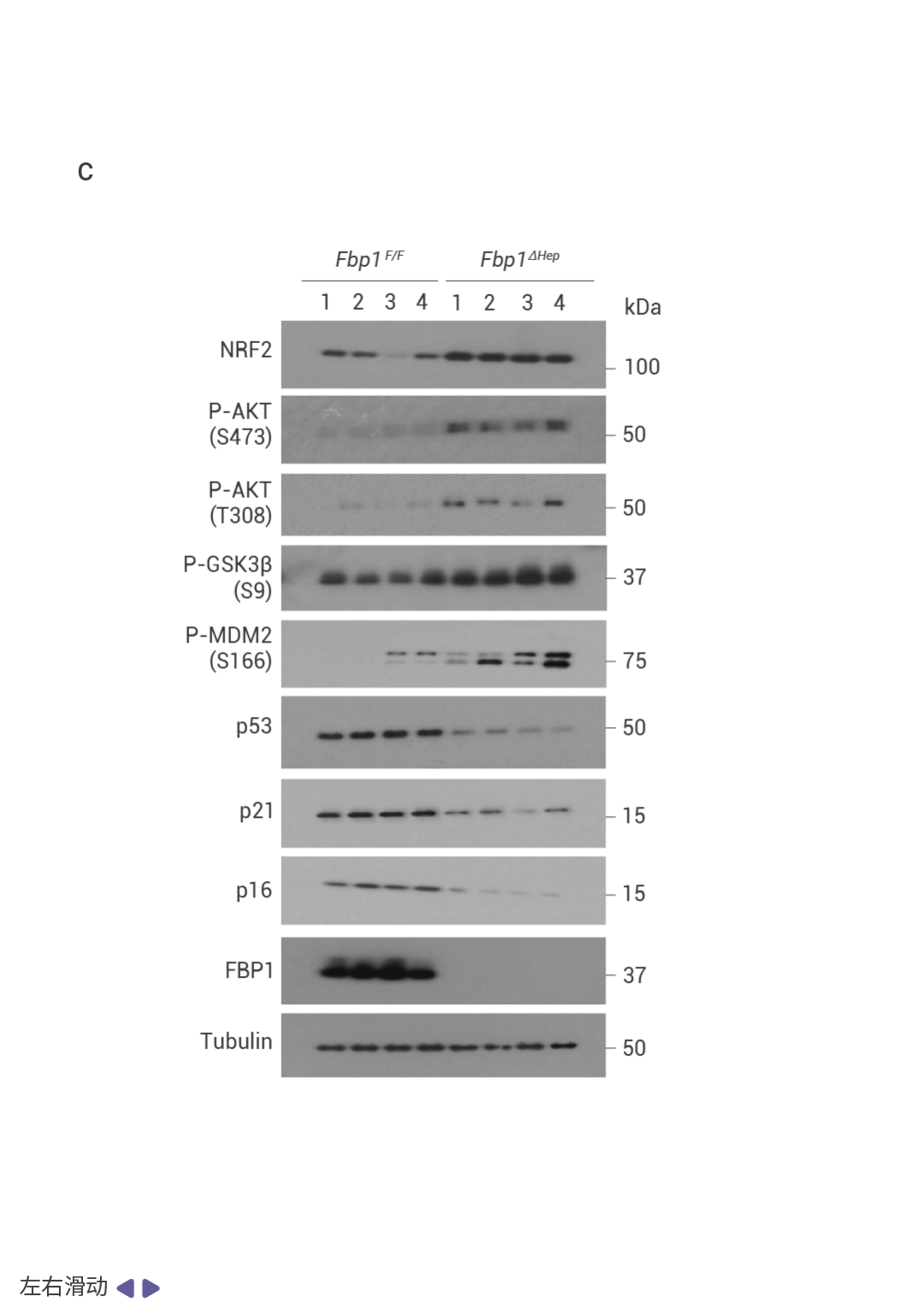
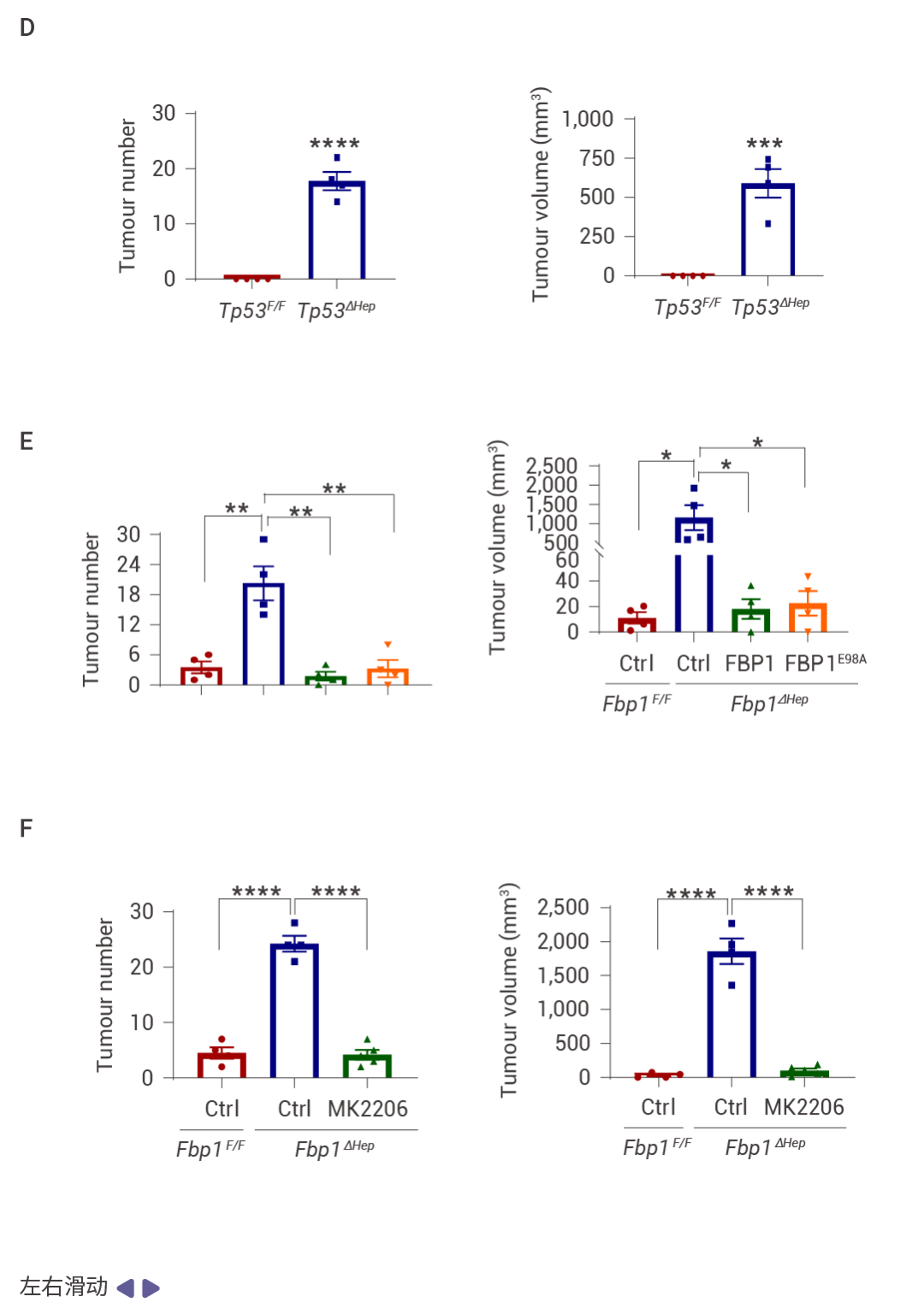
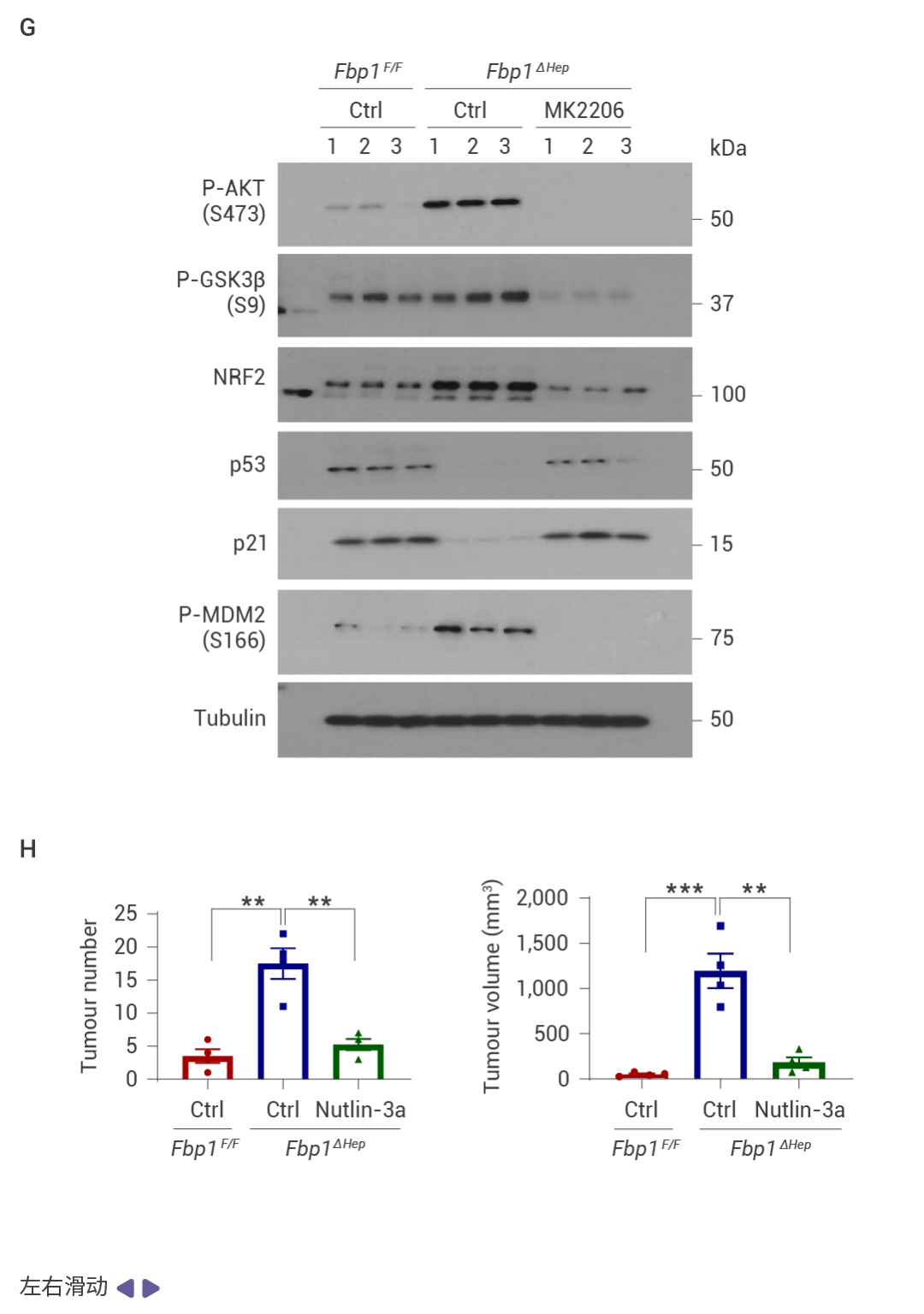 圖 3. FBP1 或 p53 缺失以及 AKT 激活促進 NRASG12V 腫瘤發生。
圖 3. FBP1 或 p53 缺失以及 AKT 激活促進 NRASG12V 腫瘤發生。
NRASG12V 在 Fbp1F/F 小鼠中幾乎不具腫瘤形成能力,但在 Fbp1ΔHep 小鼠中,它迅速誘導輕度纖維化的 HCC,這些腫瘤陽性表達 α-胎蛋白 (AFP)、CD44 和 MYC (圖 3a, b)。與 NRF2 激活一致,p53 和 p21 表達下調 (圖 3c)。肝細胞 Trp53 缺失同樣支持了快速的 NRASG12V 誘導的 HCC,同時下調 FBP1 并增強 AKT 和 NRF2 的激活 (圖 3d)。在 Fbp1F/F 和 Fbp1ΔHep 小鼠肝臟中轉導了 AAV8-Ctrl、AAV8-FBP1 和 AAV8-FBP1(E98A),兩者均顯著減少了腫瘤數量和體積 (圖 3e)。AKT 抑制劑 (AKTi) 和 nutlin-3a 也阻止了 NRASG12V 誘導的 HCC,nutlin-3a 不影響 AKT 或 NRF2,而 AKTi 阻斷了 NRF2 的激活、AKT 和 GSK3β 的磷酸化以及 MDM2 的磷酸化,同時恢復了 p53 和 p21CIP1 的表達 (圖 3f–h)。
相互調控的 NRF2–FBP1 交互作用
實驗結果表明 HCC 細胞中 FBP1 的敲除激活了 NRF2。相反,FBP1 過表達抑制了 GSK3βSer9 磷酸化,并降低了 NRF2 WT 和 NRF2 (E79Q) 的表達。NRF2 的激活導致了顯著的肝細胞增殖標記的上升,而這種變化與腫瘤發生和發展的標志密切相關。實驗還分析了 NRF2 誘導的 FBP1 降解可能是通過一種依賴于 ERK 磷酸化的途徑實現的,證明了 NRF2 通過誘導特異性生長因子,進而活化 ERK 通路,從而促進了 FBP1 的降解。這一機制表明,NRF2 可能通過降解 FBP1 這樣的腫瘤抑制因子來解除對細胞增殖和腫瘤發展過程的制約 (圖 4)。
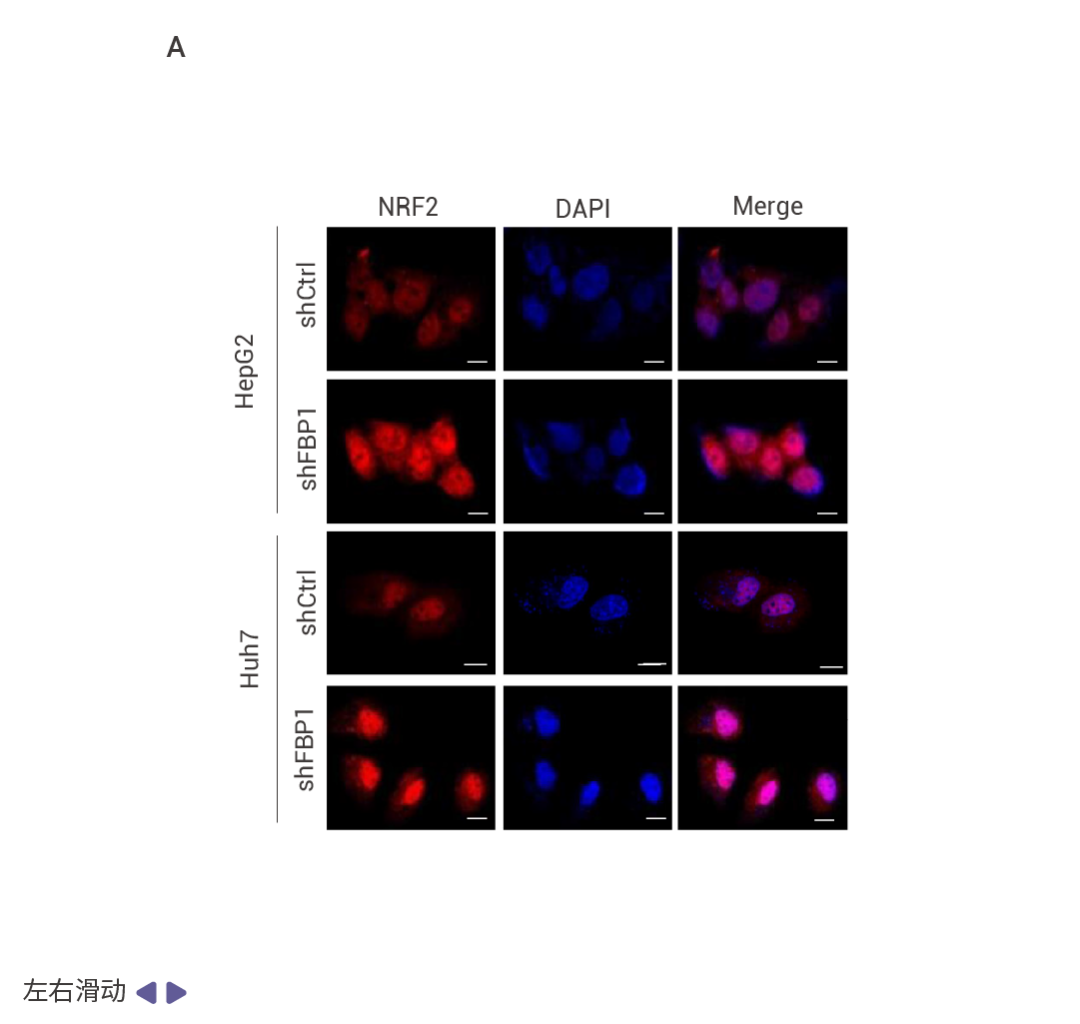
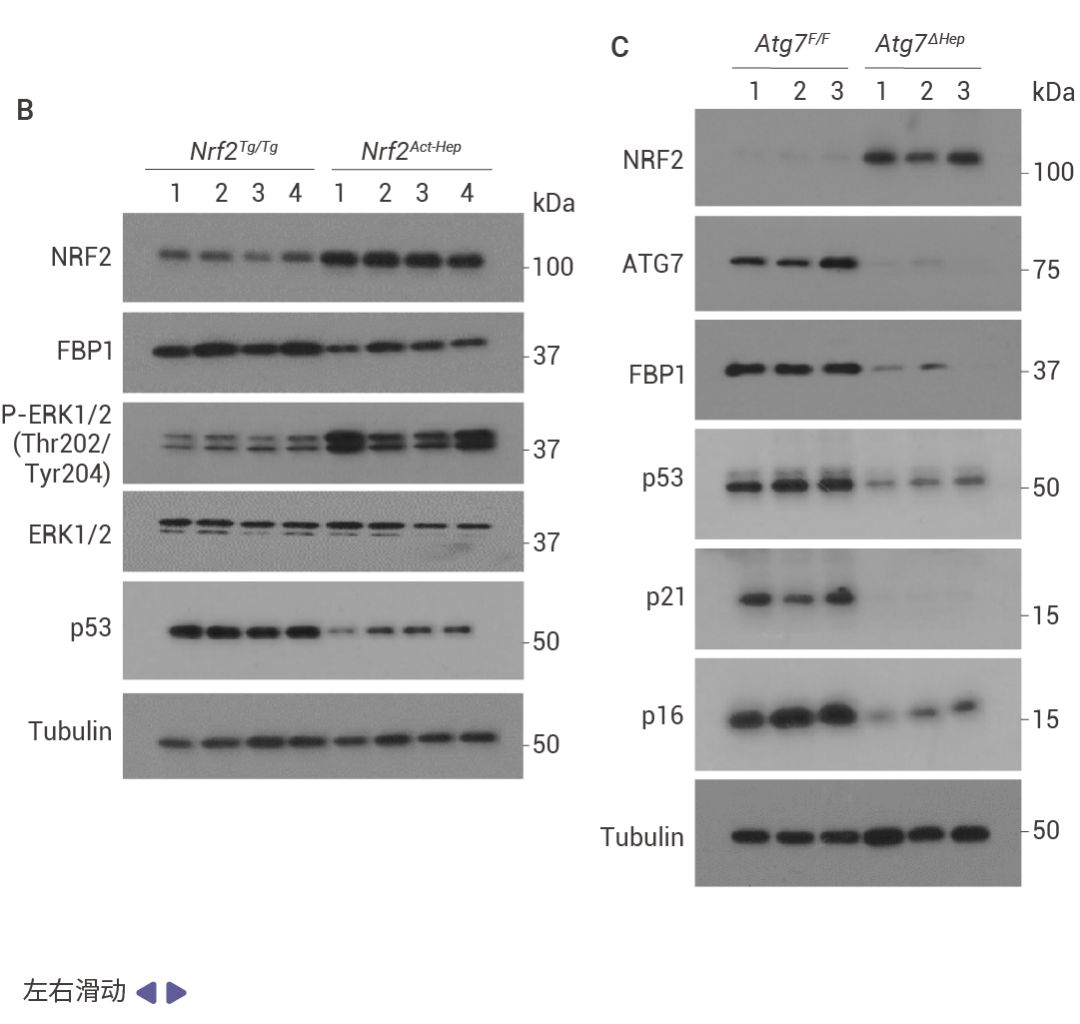
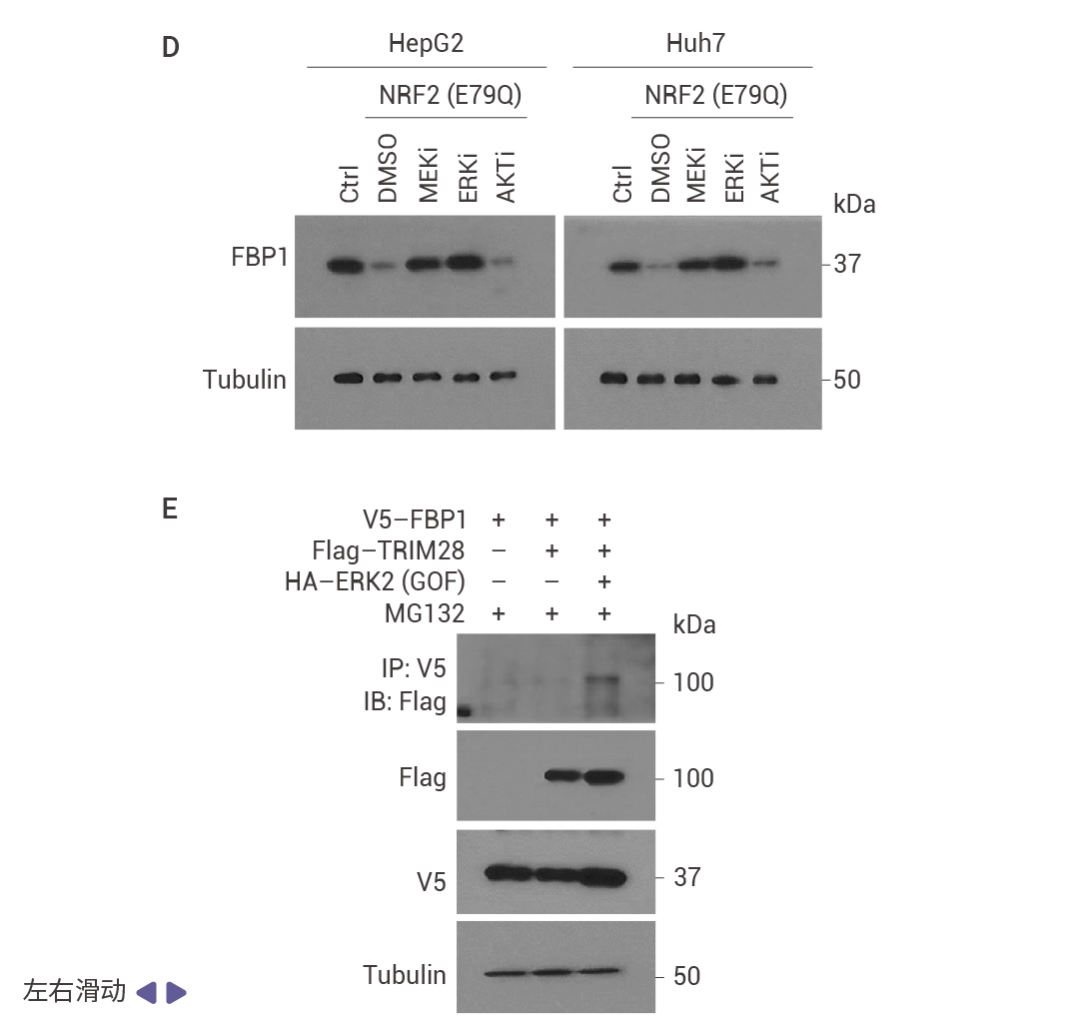
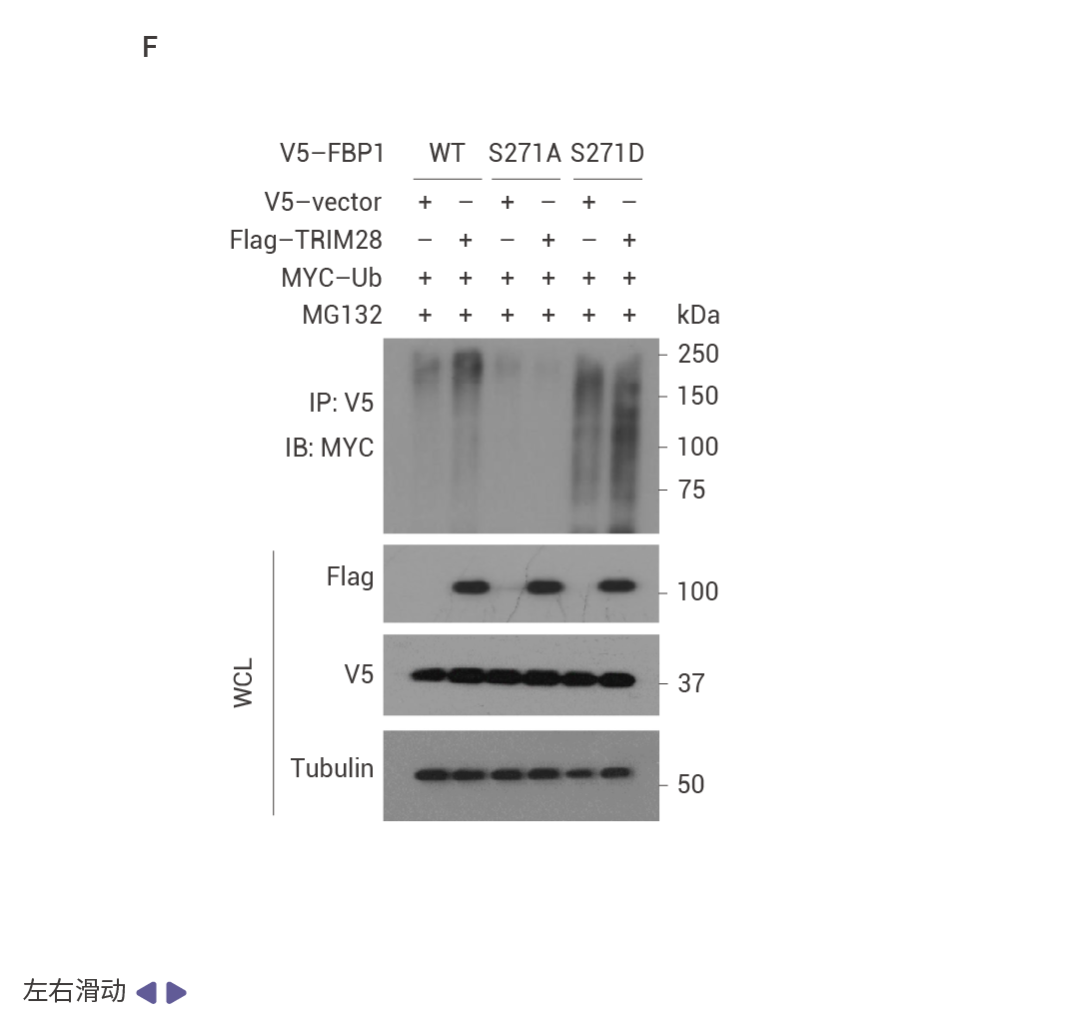
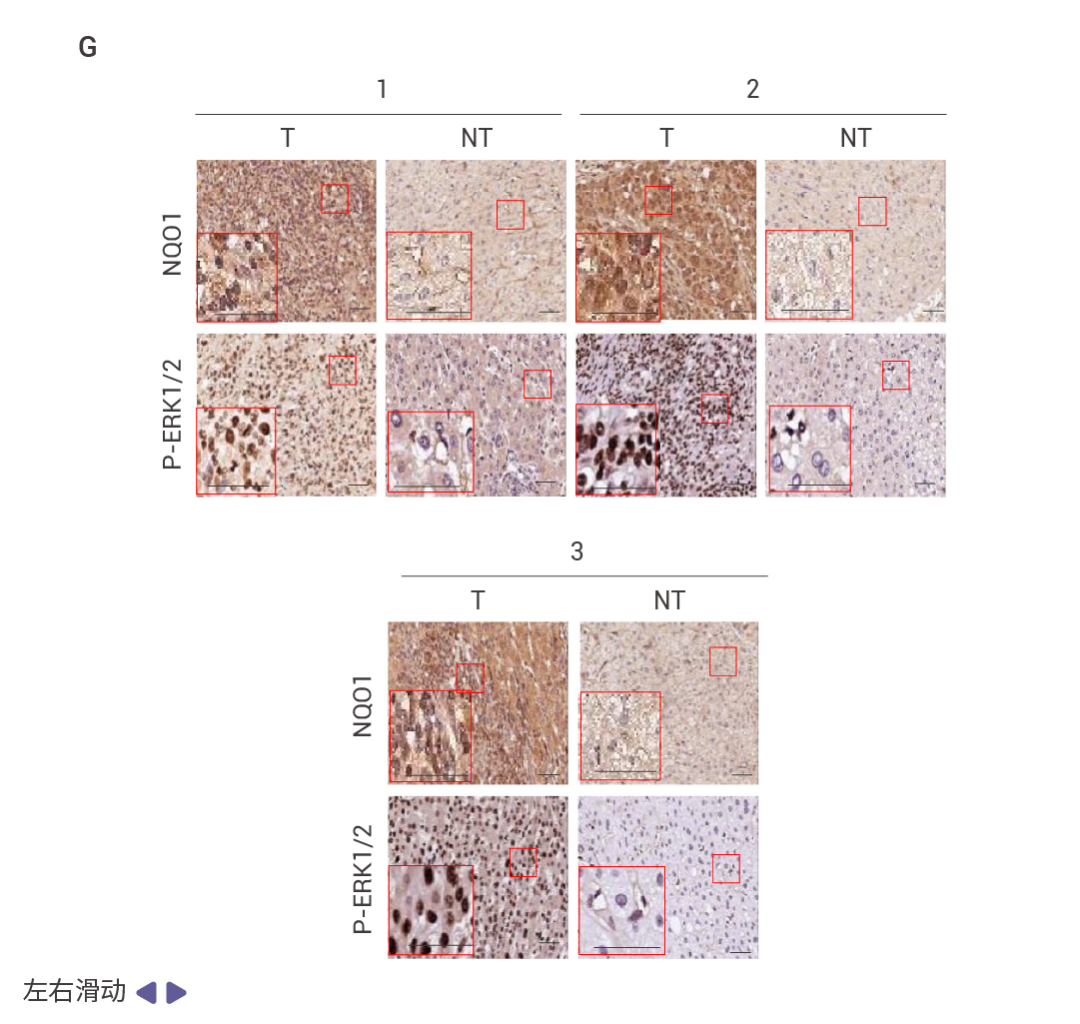 圖 4. NRF2 誘導的 FBP1 降解緩解衰老以激發腫瘤形成。
圖 4. NRF2 誘導的 FBP1 降解緩解衰老以激發腫瘤形成。
HCC 細胞中 FBP1 缺失激活了 NRF2 (圖 4a)。在 Nrf2 Act-Hep 小鼠中特異性表達 NRF2 (E79Q) 會下調 FBP1 和 p53 (圖 4b)。ATG7 缺失通過 p62 積累激活 NRF2 并增強 HCC 發展,這導致了更為顯著的 FBP1 下調 (圖 4c)。HCC 細胞中表達 NRF2 (E79Q) 加速了 FBP1 的周轉 (圖 4d),FBP1 有兩個潛在的 ERK 磷酸化受體,包括 Ser271,其中用丙氨酸替換可阻止 ERK 誘導的磷酸化及其與 FBP1 相關的泛素連接酶 TRIM28 的招募 (圖 4e)。ERK2 沉默可阻止 TRIM28 誘導的 FBP1 泛素化,而磷酸模擬突變 S271D 則增強了這一過程 (圖 4f)。FBP1 低表達的人類 HCC 還顯示出 NRF2、NQO1 和 ERK 磷酸化的上調 (圖 4g)。
NRF2–FBP1 的逆轉錄組調控
RNA-seq 分析顯示,在 Fbp1ΔHep 與 Fbp1F/F 小鼠肝臟中上調的差異表達基因中,有超過三分之一是 NRF2 誘導的,包括支持細胞增殖的脂肪酸、硫化合物、小分子、核苷、核苷酸、核糖核苷酸和氨基酸代謝相關基因。在 FBP1 缺失后下調的差異表達基因中,有四分之一與核糖核蛋白生成、非編碼 RNA 和 mRNA 代謝、肌動蛋白絲組織、細胞衰老、p53 信號傳導和凋亡調控相關,這些基因也在 Nrf2Act-Hep 小鼠肝臟中被抑制。在對照肝臟中,NRASG12V 誘導了與衰老相關的 p53 調控基因,并抑制了細胞周期基因。人類 CPTAC 數據庫顯示了類似的 FBP1 對細胞衰老和細胞周期蛋白的影響,證實了 NRF2 和 FBP1 逆向調控了一種與代謝重編程相關的抗衰老程序。
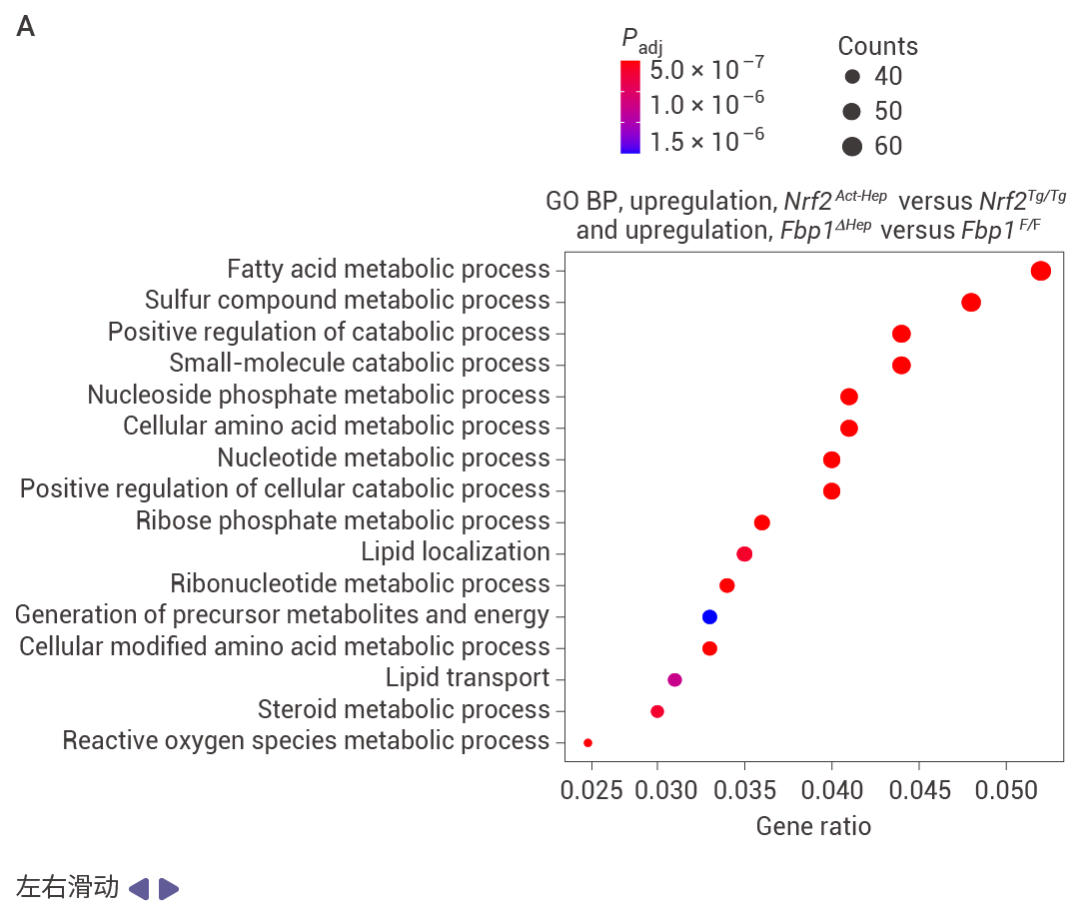
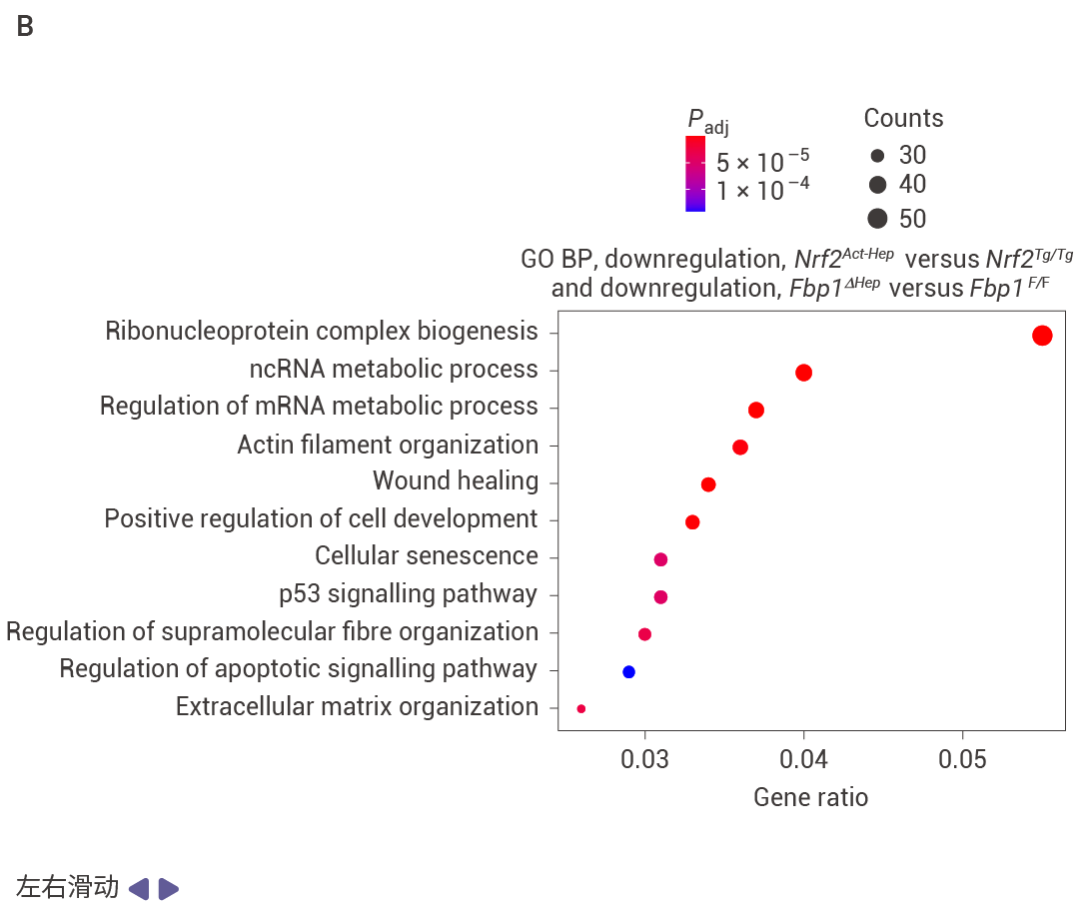
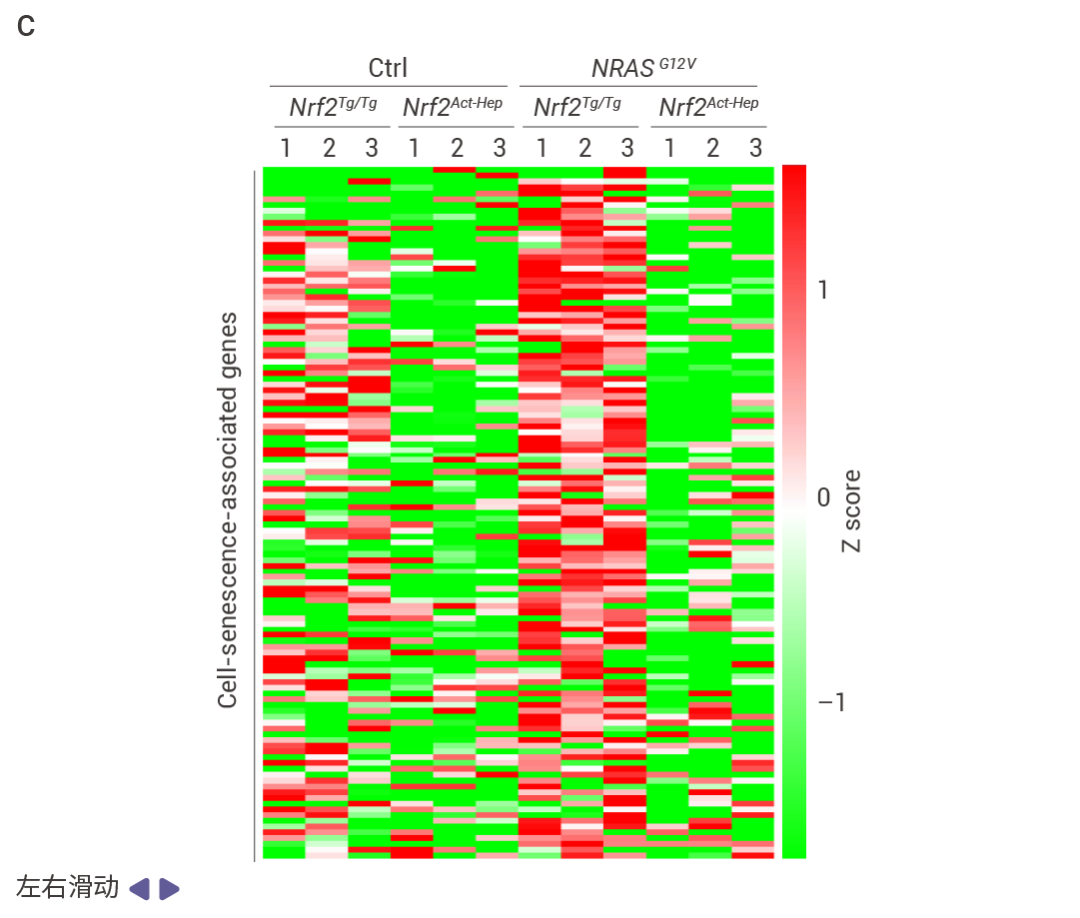
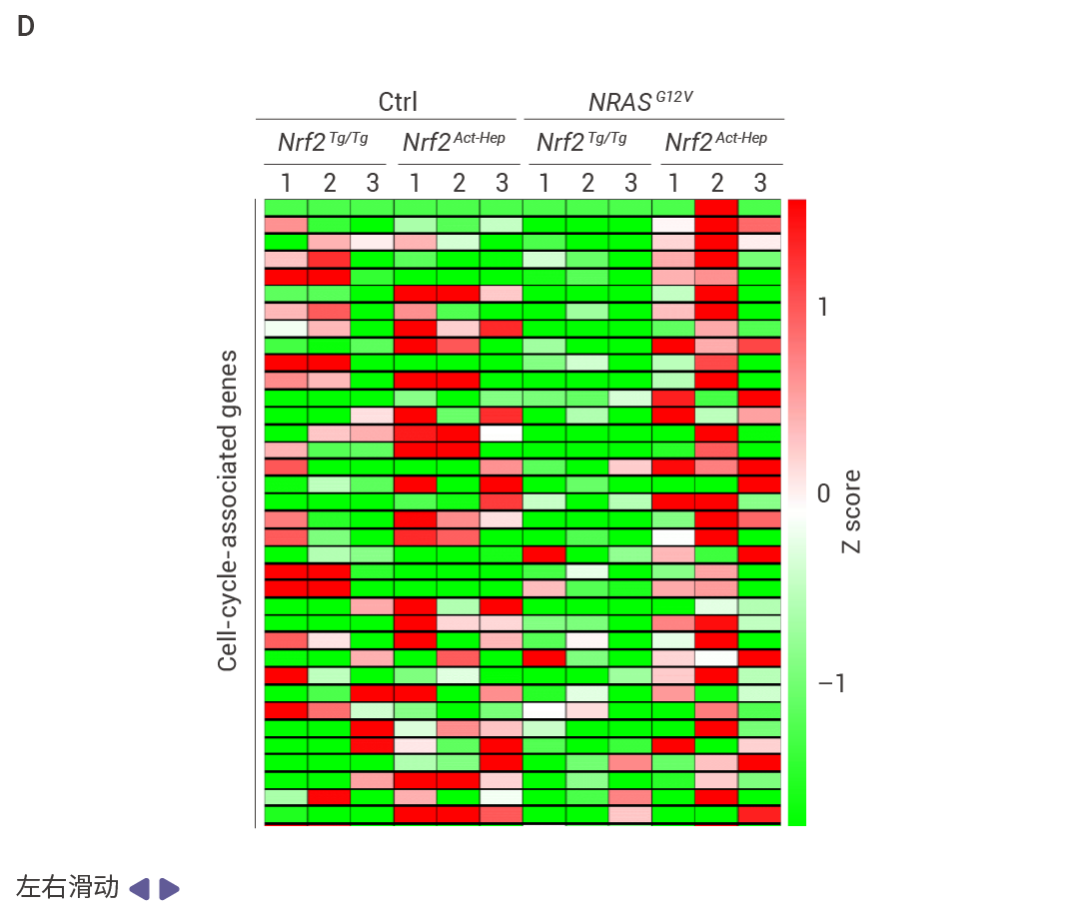
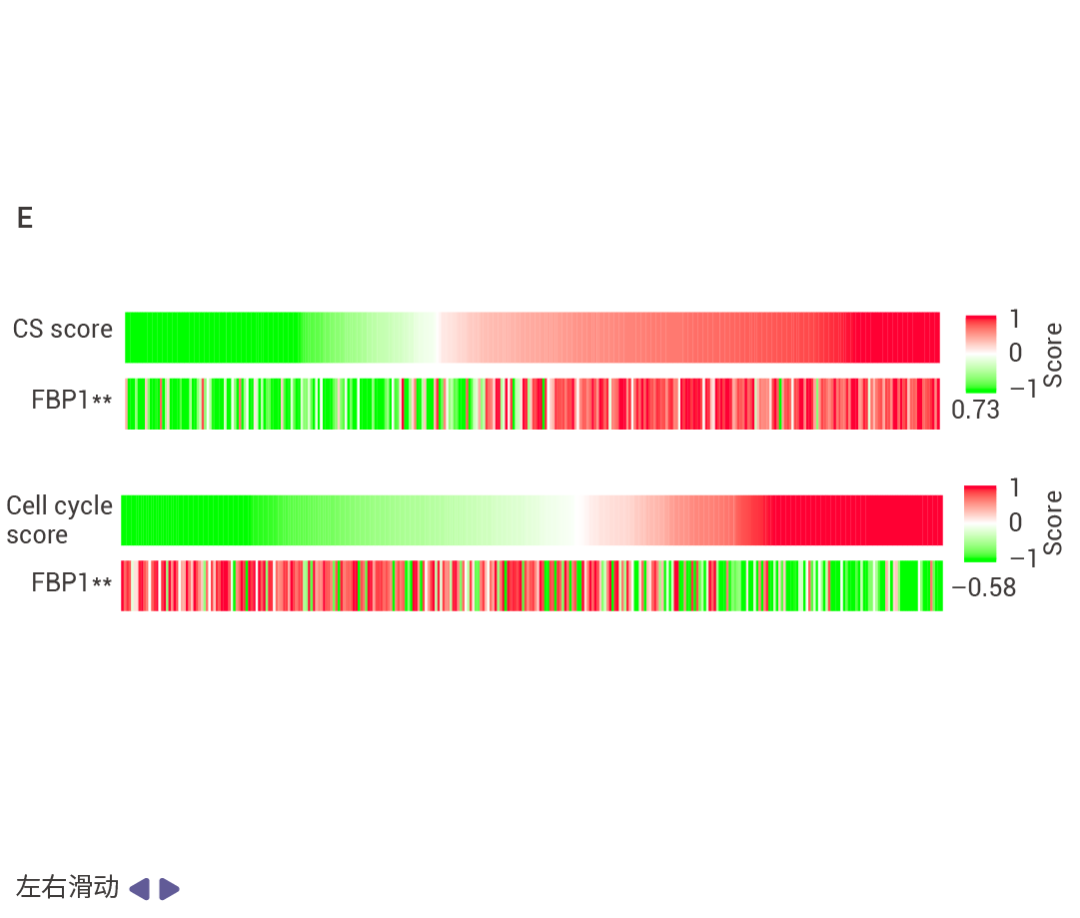 圖 5. NRF2 和 FBP1 逆向影響肝細胞轉錄組。
圖 5. NRF2 和 FBP1 逆向影響肝細胞轉錄組。
Nrf2Act-Hep 與 Nrf2Tg/Tg、Fbp1ΔHep 與 Fbp1F/F 肝臟重疊上調基因的基因本體生物學過程 (GO BP) 富集分析結果 (圖 5a)。Nrf2Act-Hep 與 Nrf2Tg/Tg 和 Fbp1ΔHep 與 Fbp1F/F 肝臟重疊下調基因的基因本體生物學過程富集分析 (圖 5b)。對照或 NRASG12V 流體動力學尾靜脈注射 (HTVI) 1 周后 Nrf2Tg/Tg 和 Nrf2Act-Hep 肝臟細胞衰老 mRNA 的熱圖數據 (圖 5c,d)。CPTAC-LIHC 數據中 FBP1 蛋白的細胞衰老 (CS) (上) 和細胞周期 (下) 評分。(圖 5e)。
NRF2 和 FBP1 對突變的相反影響
作者發現 HFD 誘導的 MUP-uPA-HCCs 含有體細胞突變。HFrD 和 HFD 誘導的 MUP-uPA-Fbp1ΔHep-HCC 的雙重測序顯示每個腫瘤平均有 500 個單堿基替換和 100 個小插入和缺失 (indel),突變模式富集了 C>T 轉換,特別是在 HFrD 誘導的 HCC 中。NRASG12V 轉導的 Nrf2Act-Hep 和 Nrf2Tg/Tg 肝臟和腫瘤的全基因組測序表明,對照肝臟幾乎不含任何突變,而 NRF2 激活的腫瘤有 1,000-2,000 個替換和 10-20 個 indels。這與人類肝細胞癌的突變模式相似。實驗結果表明,在 NRASG12V 引導下,染色體損傷觸發肝細胞衰老后,若對其施以促進 AKT 和 NRF2 通路激活的處理,這些衰老細胞會重新進入細胞周期并轉化為腫瘤。此外,綠色熒光信號主要集中在腫瘤區域,這表明衰老細胞是 HCC 發生的來源。
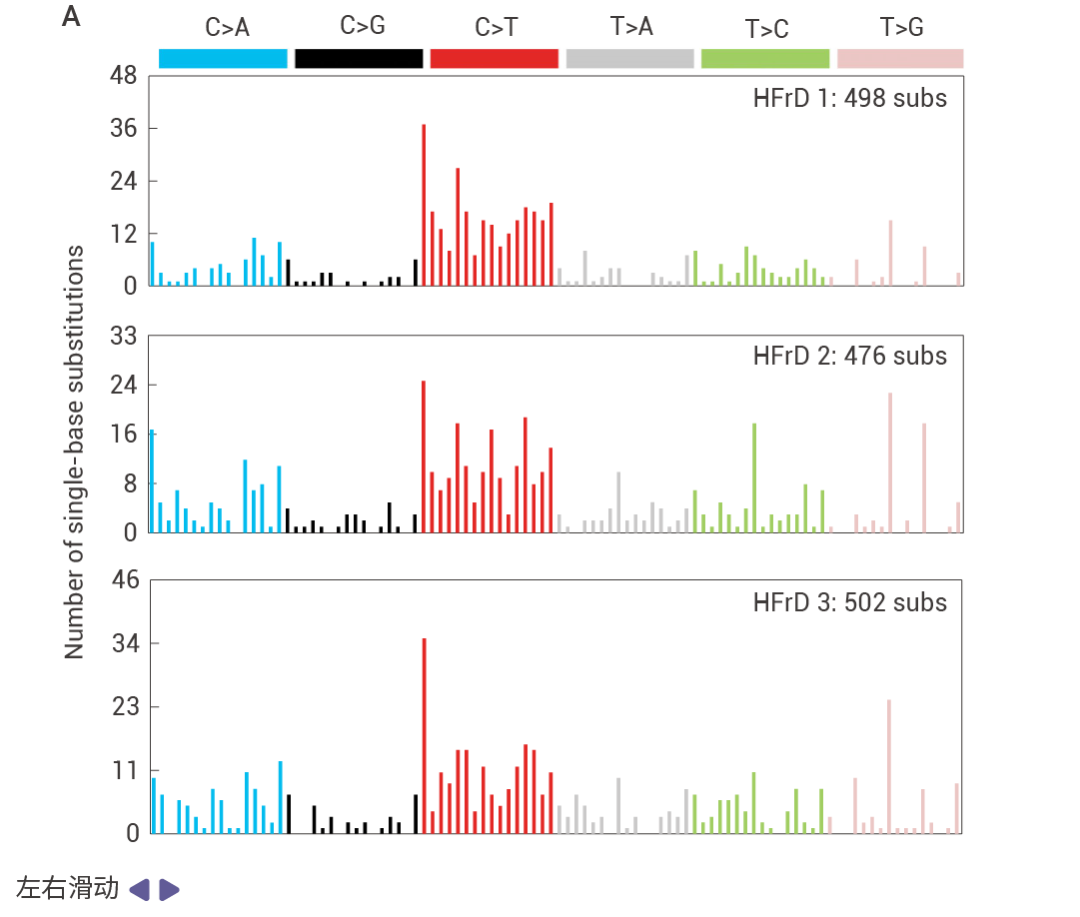
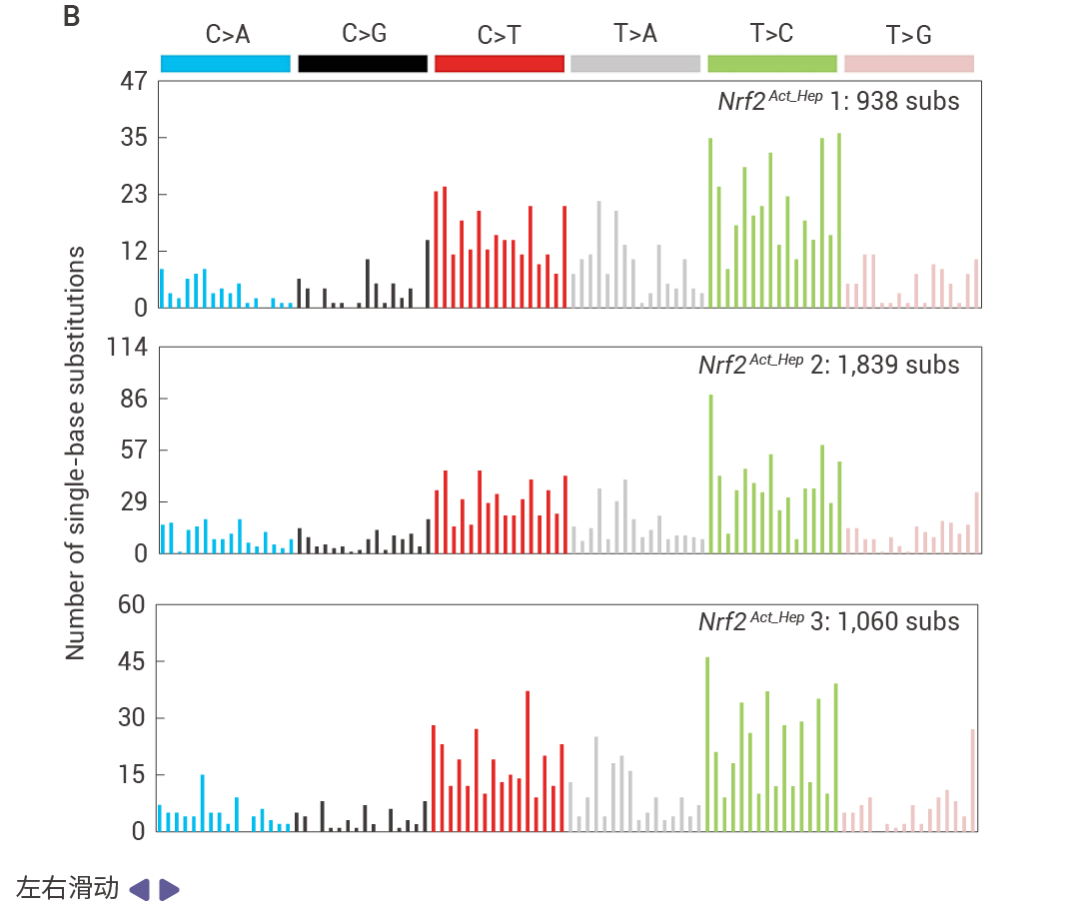
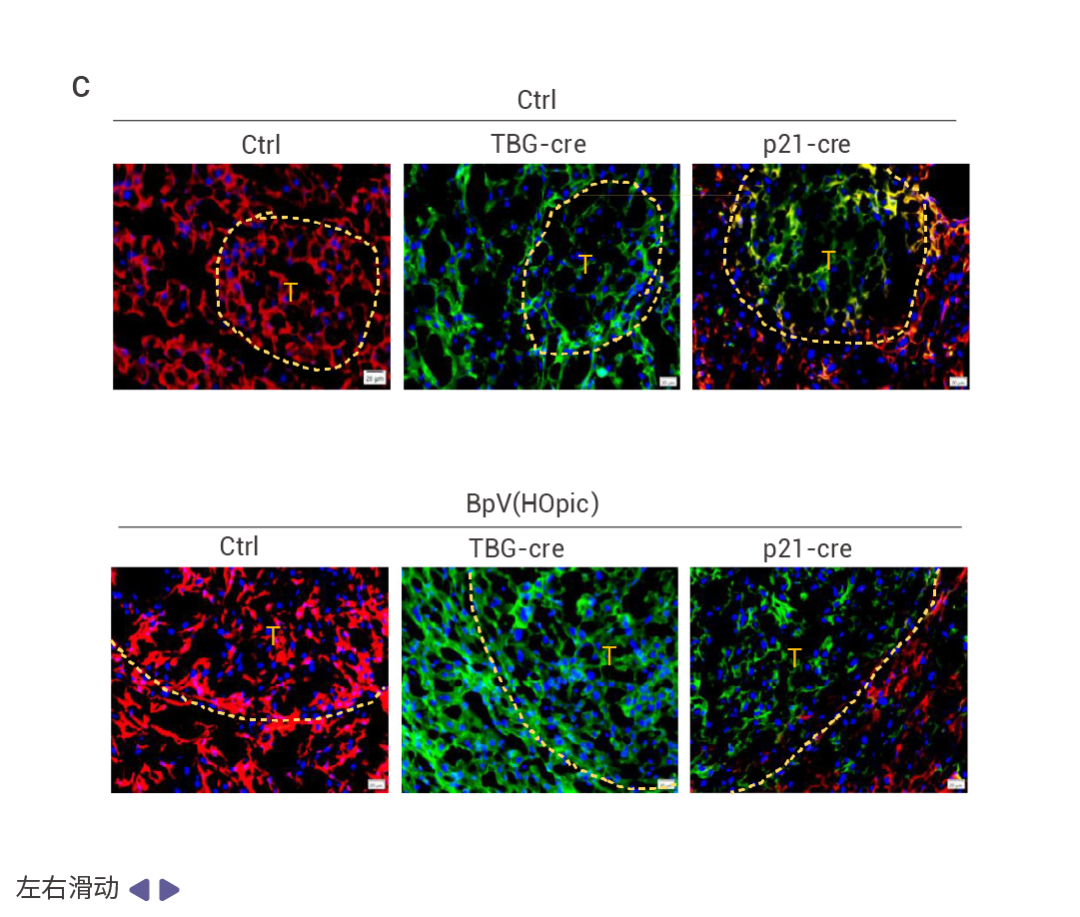 圖 6. 突變特征和 HCC 從 DNA 損傷和衰老前驅細胞中出現的過程。
圖 6. 突變特征和 HCC 從 DNA 損傷和衰老前驅細胞中出現的過程。
基于 hfrd 誘導的 MUP-uPA Fbp1ΔHep HCC 的三核苷酸的突變頻率特征 (圖 6a)。 NRASG12V 誘導的 Nrf2Act-Hep HCC 的突變頻率特征 (圖 6b)。冷凍 mT/mG 肝臟切片 DAPI 染色結果 (圖 6c)。
Section.03
小結
MASH 是如何在增加肝細胞癌風險的同時又抑制肝細胞癌的發生?該研究表明,MASH 增加了肝癌的風險,同時觸發腫瘤抑制的肝細胞衰老。通過詳細的組織學、蛋白質組學和轉錄組學分析,研究發現 MASH 肝細胞高表達 FBP1、p53 和關鍵衰老標志物,而與之相對的疾病相關肝細胞 (daHeps)、HCC 前體細胞 (HcPCs) 和確立的 HCC 則表現出 FBP1 和 p53 下調。這種下調可能是由于在代謝壓力的 HCC 前體中 NRF2 激活所致。研究揭示了 NRF2 通過誘導 ERK 活化的生長因子觸發 FBP1 的磷酸化依賴性降解,解除 AKT 活化的抑制,加速 p53 降解和衰老逆轉。這些調節相互作用傾向于促進 HcPCs 擴增,使 DNA 損傷誘導的突變得以傳播并導致腫瘤發生。研究強調了 NRF2 和 AKT 激活或 FBP1 缺失在肝細胞衰老和肝癌進展中的重要作用,并為開發有效的 DNA 損傷抑制劑以用于 MASH 和 HCC 的預防提供了理論支持。
| 產品推薦 |
| Etoposide Etoposide (VP-16; VP-16-213) (HY-13629),是一種常用的抗腫瘤化療劑。Etoposide 抑制拓撲異構酶 II (topoisomerase-II),誘導細胞周期停滯,凋亡 (apoptosis) 和自噬 (autophagy)。 |
| 5-Azacytidine 5-Azacytidine (Azacitidine; 5-AzaC; Ladakamycin) (HY-10586),是胞苷核苷類似物,特異型抑制 DNA 甲基化。5-Azacytidine 誘導細胞自噬 (autophagy)。 |
| BpV (HOpic) BpV (HOpic) (HY-128693),是一種有效的和選擇性的 PTEN 抑制劑,IC50 值為 14 nM。Nanocarrier-BpV (HOpic) 具有神經保護作用。 |
| NK252 NK-252 (HY-19734),是一種潛在的 Nrf2 激活劑,具有很好的 Nrf2 活化能力。 |

[1] Gu, L., Zhu, Y., Nandi, S.P. et al. FBP1 controls liver cancer evolution from senescent MASH hepatocytes. Nature 637, 461–469 (2025).
[2] Huang DQ, et al. Global epidemiology of NAFLD-related HCC: trends, predictions, risk factors and prevention. Nat Rev Gastroenterol Hepatol. 2021 April ; 18(4): 223–238.


本站“ABIO生物試劑品牌網”圖片文字來自互聯網
如果有侵權請聯系微信: nanhu9181 處理,感謝~