睡眠壓力的線粒體起源及大腦中分子變化與睡眠需求的關聯_abio生物試劑品牌網
線粒體:哈?人!關掉你的網抑云,我要裂了......
Section.01
人為什么會睡覺?
你有沒有想過,我們為什么會睡覺? 我指的是......不受控的,打瞌睡。無論你憑借意志陪月亮熬到凌晨幾點,終有一個瞬間閉眼投降。緊跟著的便是熬夜過后身體的種種不良反應,讓你感覺身體被掏空,下一秒就要見到老天奶。。。
其實從進化的角度看,睡眠并不是件好事。因為如果生物體長時間無法感知和與環境互動,可能會增加被捕食的風險。但神奇的是! 從秀麗隱桿線蟲的簡單神經系統,一直到人類大腦,睡眠從未被進化掉 ?! [1] 。
于是,科學家們將睡眠稱之為一種非常特殊的現象。加之睡眠不足會帶來認知能力受損、代謝功能障礙、患病風險增加,極端情況下甚至會導致死亡等有害影響。So,睡眠,既是動物界最常見的行為之一,也是自然界最大的謎團之一。
你可能要說了,存在即合理嘛~ 哎嘿!兄弟,它不僅合理,還有規律——生物鐘。
生物鐘,又稱晝夜節律 (Circadian rhythm):是一種生理現象,以內源性、持續的,呈現以約 24 小時為周期的變動。
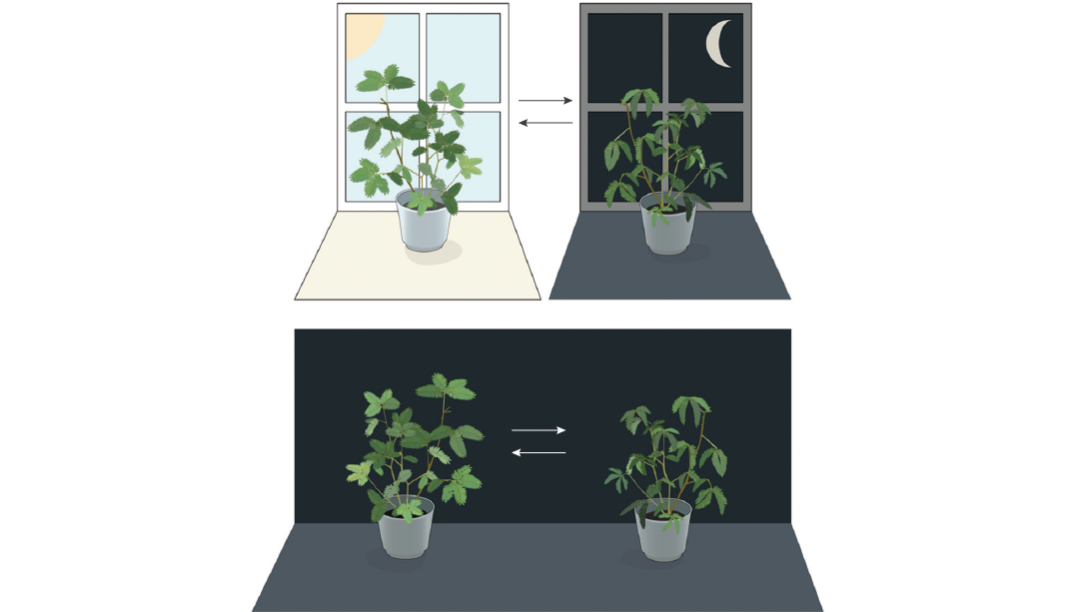 圖 1. 含羞草的葉片白天張開,黃昏時閉合。且當含羞草處于持續黑暗中時,其葉片仍會維持正常的晝夜節律
[2]
。
圖 1. 含羞草的葉片白天張開,黃昏時閉合。且當含羞草處于持續黑暗中時,其葉片仍會維持正常的晝夜節律
[2]
。
早在 18 世紀,天文學家就發現植物具有內源性的生物節律。為了了解這種內源性晝夜節律的機制,科學家做了諸多研究。直到多年后,3 位美國科學家杰弗理?霍爾 (Jeffrey C. Hall)、邁克爾?羅斯巴希 (Michael Rosbash)、邁克爾?楊 (Michael W. Young) 揭示了果蠅生物鐘的調控機制,也因此榮獲 2017 年的諾貝爾生理學或醫學獎。
科學家成功克隆了 "周期" 基因 period,period 基因的編碼蛋白 PER 夜間積累,白天降解,其蛋白水平的 24 h 周期性波動與晝夜節律同步。TIM 蛋白可以與 PER 蛋白結合,攜帶后者進入細胞核,通過抑制 period 基因活性來完成負反饋調控,減少自身蛋白的合成。此外,基因 doubletime 編碼的蛋白 DBT 可以延緩 PER 蛋白的積累,使后者的震蕩頻率基本吻合 24 h 周期 (圖 2) 。
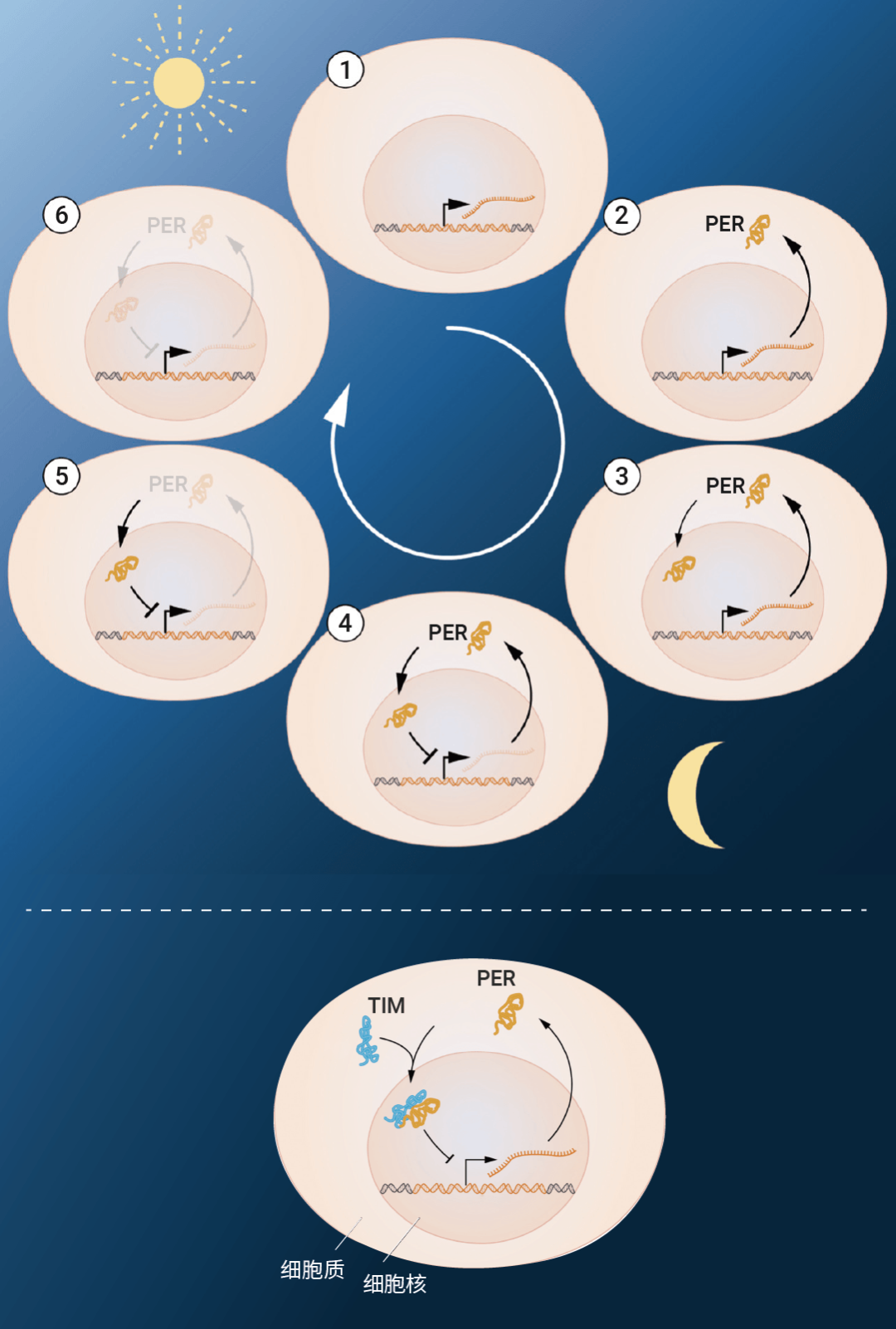 圖 2. period 基因負反饋調控環路
[2]
。
圖 2. period 基因負反饋調控環路
[2]
。
2017 年的諾貝爾生理學或醫學獎也讓果蠅這一卓爾不凡的小動物再次身顯名揚,這也是第五座因果蠅而獲得的諾獎。后續小 M 會位大家專門出一期關于實驗中常見模式生物的文章,小伙伴們可以淺淺關注一下~
言歸正傳!果蠅,作為研究睡眠相當有用的動物模型 [3] 。研究發現,在其睡眠調節神經回路中,背側扇形體 (dorsal fan-shaped body, dFB/dFNBs) 是主要的促進睡眠區域 [4] 。更為神奇的是,近期 Nature 發表的研究性論文發現,睡眠與線粒體緊密相關 ——當在背側扇狀體 (dFBNs) 中檢測到過多的線粒體損傷時,就會觸發睡眠。
Section.02
Nature 發文:
線粒體起源的睡眠壓力
2025 年 7 月 16 日,Nature 發表標題為“ Mitochondrial origins of the pressure to sleep ”的研究性論文,該論文研究了睡眠壓力 (sleep pressure) 的線粒體起源,探討了大腦中分子變化與睡眠需求的關聯[5]。
線粒體基因表達上調
首先,研究人員通過分析休息和睡眠剝奪的果蠅的單細胞轉錄組,發現在睡眠剝奪后,上調的轉錄本主要在投射到背側扇狀體 (dFBNs) 的睡眠控制神經元中表達,且 在 dFBN 中上調的轉錄本,幾乎只編碼參與線粒體呼吸和 ATP 合成的蛋白質 (圖 3a) 。
如圖所示,通過對 12 小時夜間睡眠剝奪后 dFBN 中水平發生變化的 122 個轉錄本進行基因本體分析,發現睡眠剝奪導致編碼電子傳遞復合物 I-IV、ATP 合酶 (復合物 V) 、ATP-ADP 載體 sesB 和三羧酸循環酶 (檸檬酸合酶 kdn、琥珀酸脫氫酶 B 亞基和蘋果酸脫氫酶 Men-b) 成分的轉錄本選擇性上調,而參與突觸組裝、突觸囊泡釋放和突觸前穩態可塑性的基因產物則選擇性下調。
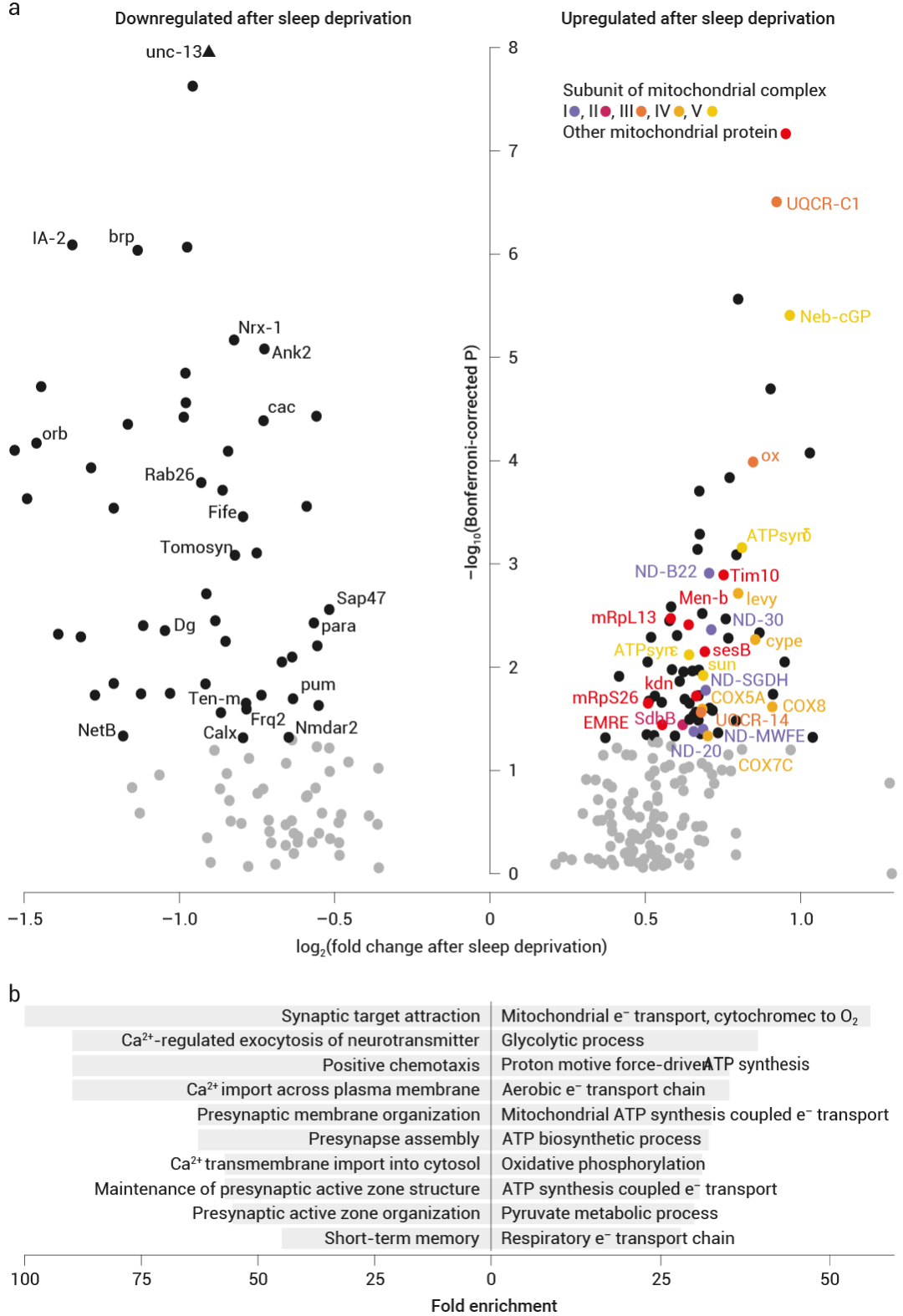 圖 3. 線粒體成分在 dFBN 對睡眠剝奪的轉錄反應中的突出作用
[5]
。
圖 3. 線粒體成分在 dFBN 對睡眠剝奪的轉錄反應中的突出作用
[5]
。
a. dFBN 中睡眠史依賴性基因表達變化的火山圖。Bonferroni 校正的 P<?0.05 的信號以黑色表示;標簽標識定位于突觸或線粒體的蛋白質產物;顏色表示線粒體呼吸復合體的亞基。b. 差異表達的 dFBN 基因集中前十個下調(左)和上調(右)的“生物過程”基因本體術語的富集。
睡眠剝奪 — 線粒體: 不睡么? 我要 "裂" 了
作者用基質定位的綠色熒光蛋白 (mito-GFP) 標記了 dFBN 的線粒體,并在一系列獨立的實驗中,用共聚焦激光掃描顯微鏡 (CLSM) 或光學光子重排顯微鏡 (OPRM) 對神經元的樹突區進行成像。發現 睡眠剝奪后的基因表達變化伴隨著線粒體碎片化、線粒體自噬增強以及線粒體與內質網之間接觸數量的增加,恢復睡眠后,形態變化是可逆的。
首先,在線粒體形態上,研究發現無論是機械攪拌還是人為提高多巴胺水平所致的睡眠剝奪,都會導致 dFBN s 中線粒體的尺寸縮小、伸長率降低、分支減少 (圖 4a-b) 。并導致動力蛋白相關蛋白 1 (Drp1) (外膜的關鍵裂變動力蛋白) 從細胞質移位到線粒體表面 (圖 4c) ,OPRM 檢測到線粒體數量的伴隨增加 (圖 4b) ,表明線粒體發生分裂。同時,睡眠剝奪的果蠅的 dFBN 中接觸位點計數較高,也就是說睡眠剝奪導致 dFBN 中線粒體-內質網接觸更加豐富,從而形成補充過氧化脂質的通道 (圖 4d) 。同時,mito-QC (一種檢測線粒體進入酸性自噬溶酶體的比例傳感器) 則報告了線粒體自噬增強 (圖 4e) 。而在隨后的恢復睡眠期間,線粒體的增殖和融合導致它們的體積、形狀和分支長度反彈至基線值以上 (圖 4a-b) 。 這表明線粒體損傷可能會引發睡眠沖動。
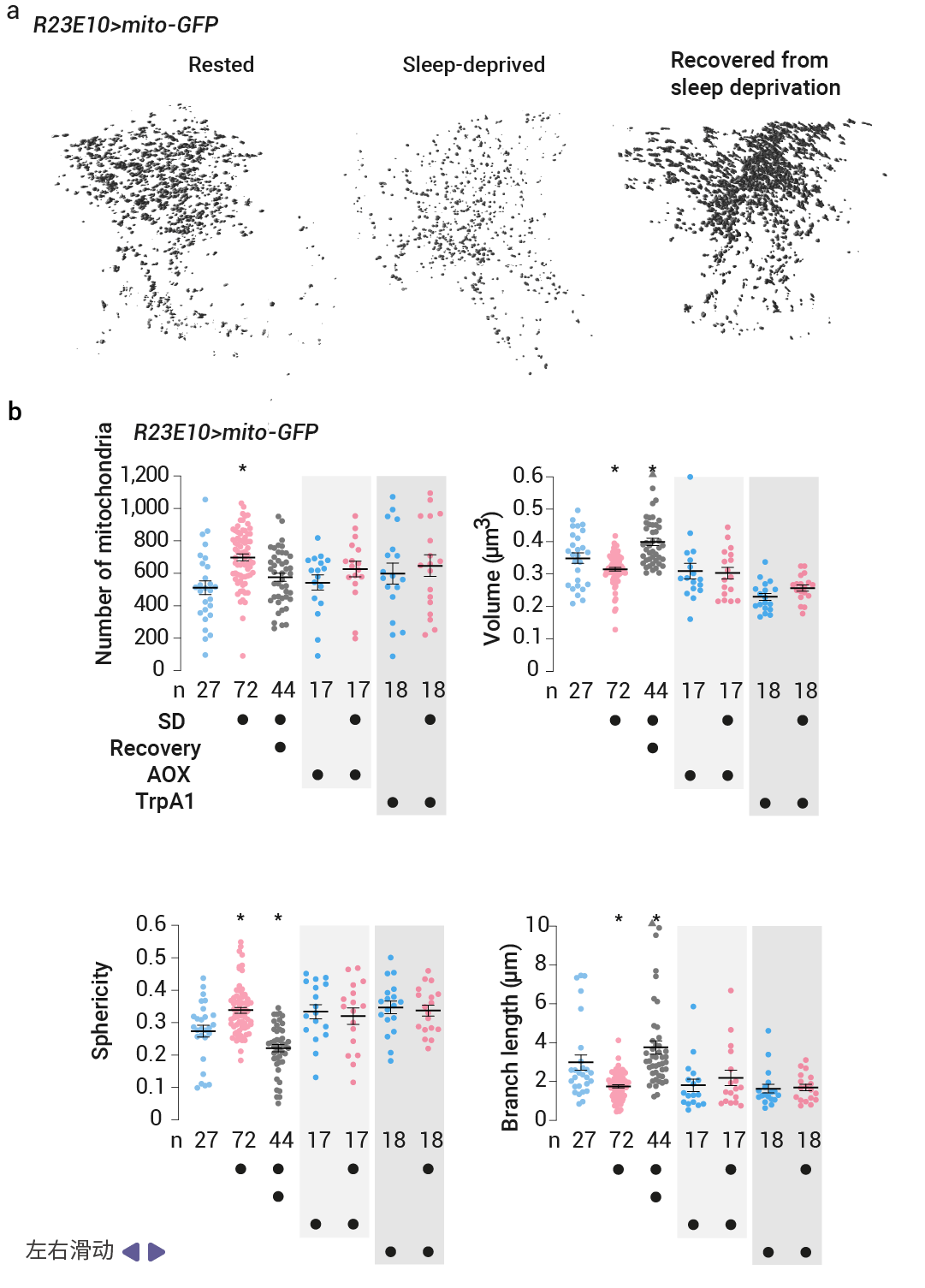
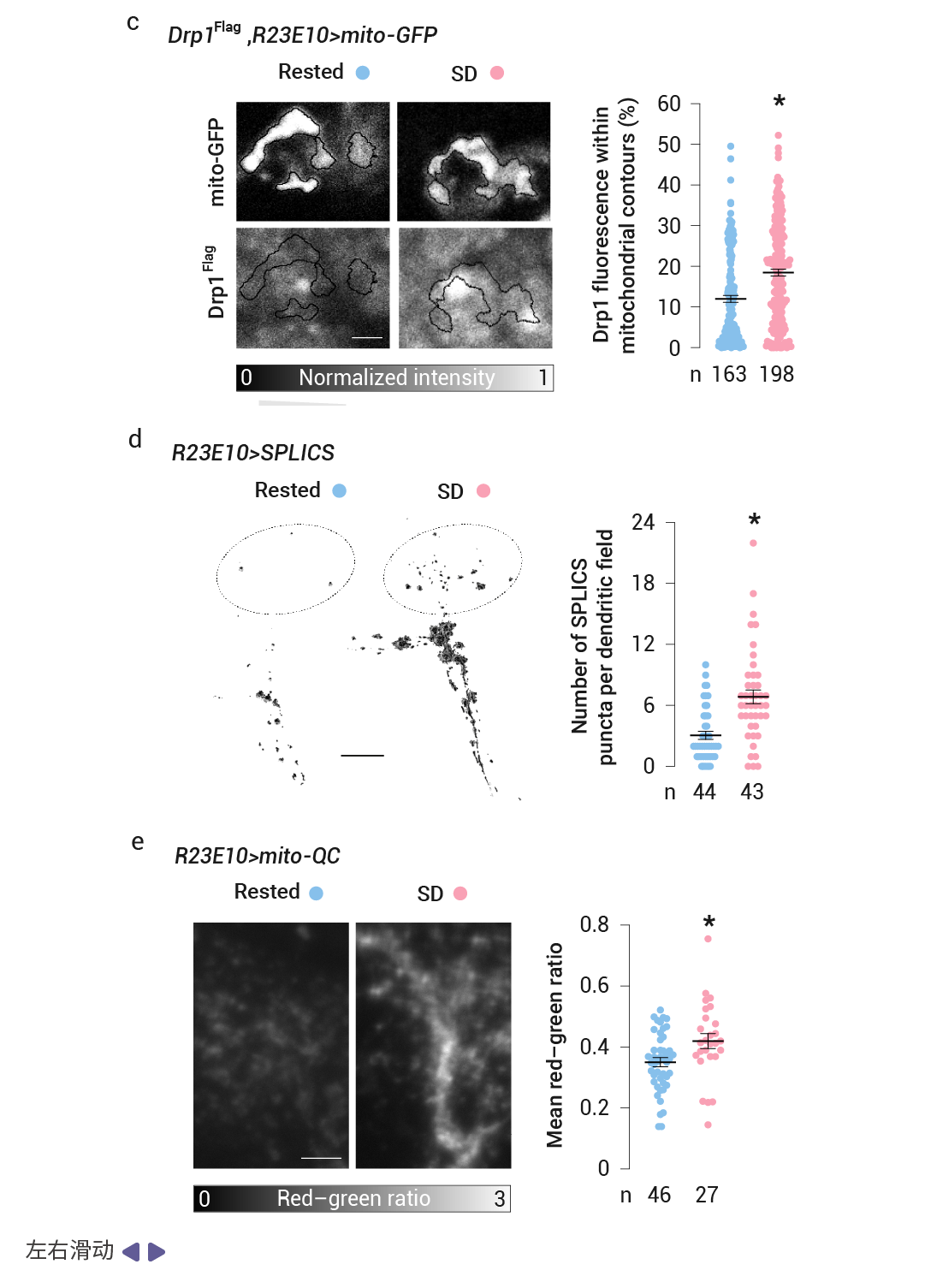
圖 4. 睡眠改變線粒體的動態變化 [5] 。
a-b. 休息的蒼蠅、睡眠不足的蒼蠅和睡眠不足 24 小時后恢復的蒼蠅的 dFBN 樹突的 OPRM 圖像堆棧中自動檢測到的線粒體的體積渲染 (a) 和形態參數 (b)。c. Drp1 募集。通過表達 R23E10-GAL4 驅動的線粒體-GFP(頂部)和來自內源性基因座的 Drp1 Flag (底部)的果蠅的dFBN 體細胞的單共聚焦圖像平面。睡眠剝奪會增加自動檢測到的線粒體輪廓內的細胞抗 Flag 熒光的百分比。d. 線粒體-內質網接觸。e. 線粒體自噬。
此外,利用基因編碼的 ATP 傳感器 iATPSnFR 和 ATeam 進行的測量表明,一夜睡眠剝奪后,dFBN 中的 ATP 濃度比靜息狀態下高出約 1.2 倍。而當 dFBN 本身受到刺激 (模擬睡眠) 時,ATP 濃度會降至基線以下,這是由于 dFBN 去極化會增加 Na + -K + 泵的 ATP 消耗,從而減少電子向 ROS 的轉移 (圖 5) ,這保留了其線粒體的形態。
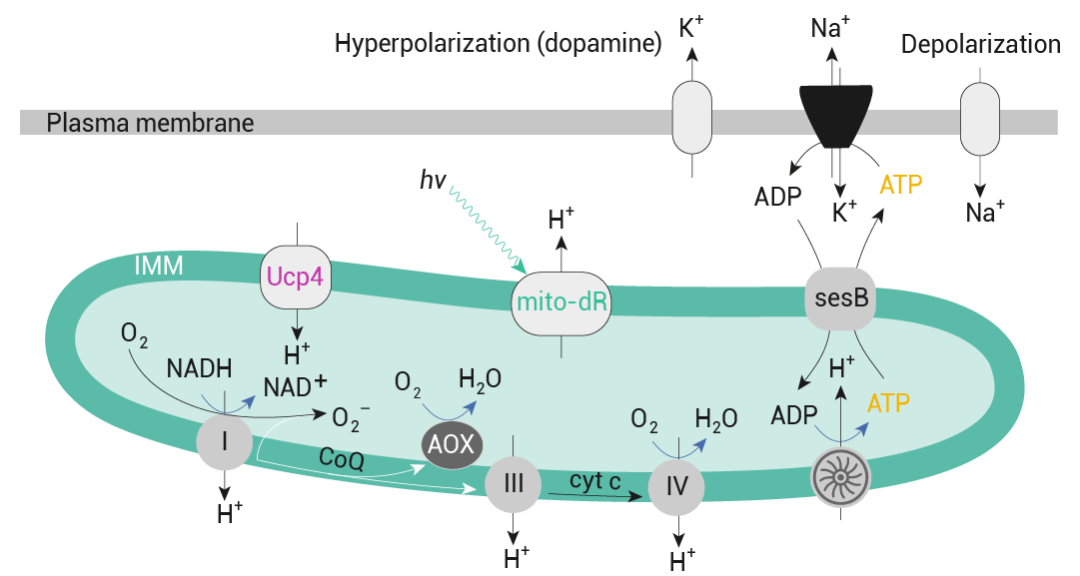 圖 5. dFBN 通過將誘導睡眠的脈沖放電與線粒體呼吸相結合來調節睡眠
[5]
。
圖 5. dFBN 通過將誘導睡眠的脈沖放電與線粒體呼吸相結合來調節睡眠
[5]
。
質子泵復合物 I、III 和 IV 將電子從 NADH 轉移到 O 2 (通過中間體 CoQ 和細胞色素 c (cyt c)) 的能量轉化為跨 IMM 的質子電化學梯度 ?p。Ucp4 放電,而 mito-dR 的光照 (hν) 使 IMM 充電。擠出的質子返回基質使 ATP 合酶的葉片旋轉并產生 ATP,ATP 通過 sesB 離開基質,與細胞質 ADP 交換。神經元 ATP 消耗依賴于活動,部分原因是質膜 Na + -K + ATPase 必須恢復因動作和興奮性突觸電流而消散的離子梯度。CoQ 電子供應過量 (相對于 ATP 需求)會增加復合物 I 和 III 處 O 2 單電子還原為 O 2 ? 的風險。AOX 可降低這種風險。
線粒體:裂變—睡的少,融合—叫不醒
隨后,作者人為地人為控制了線粒體的裂變和融合。因為如果線粒體裂變和融合之間平衡的變化是反饋機制的一部分。那么在 dFBN 中實驗性地誘導這些穩態反應應該會改變睡眠的設定點: 線粒體的分裂預計會減少睡眠時間和深度,而反之線粒體融合預計會增加睡眠時長和深度。
于是,作者對在線粒體動力學中具有中心調控作用的三種 GTP 酶進行了實驗控制 (圖 4a) :(1) 裂變動力蛋白 Drp1,以及內外線粒體膜的整合蛋白 (2) 視神經萎縮 1 (Opa1) 和 (3) 線粒體融合蛋白子 (Marf) (它們在順式和反式的聚合使得相應的膜融合) 。
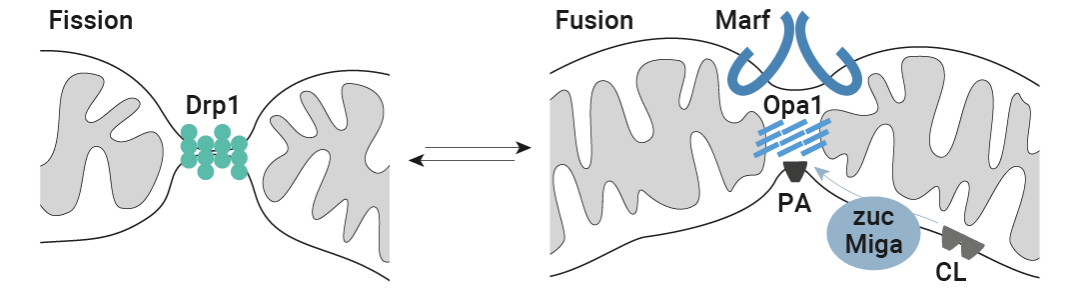 圖 6. 線粒體裂變 (綠色) 和融合機制 (藍色)
[5]
。
圖 6. 線粒體裂變 (綠色) 和融合機制 (藍色)
[5]
。
線粒體裂變 (綠色) 和融合機制 (藍色) 包括 Drp1、外膜蛋白和內膜蛋白 Marf 和 Opa1,以及線粒體 PLD zuc,它從心磷脂 (CL) 中釋放磷脂酸 (PA)。Miga 刺激 zuc 活性和/或從其他膜提供磷脂酸。
通過過表達 Drp1 或 RNAI 介導的 Opa1 減少 (在較小程度上也包括 Marf) 來使 dFBN 線粒體碎片化,減少了睡眠 (圖 7a-b) 。消除對睡眠剝奪的穩態反應 (圖 7c) ,并且無論睡眠史如何,均降低了 dFBN 中的 ATP 濃度。
將平衡傾向于線粒體融合則產生了相反的效果:在 dFBN 中敲低 Drp1 或過表達 Opa1 加 Marf——或者單獨過表達 Opa1,但不包括單獨過表達 Marf——增加了基線及反彈睡眠 (圖 7a-c) ,并提高了覺醒閾值,且不會引起過表達偽影或明顯的發育缺陷。
同時,當這些干預針對投射神經元或 Kenyon 細胞時,沒有改變睡眠。報道顯示,在泛神經元或膠質細胞 RNAi 敲低 Drp1 或 Marf 基因后,睡眠喪失現象普遍存在。在 dFBN 中,Drp1 或 Marf 基因的表達會雙向改變睡眠 (圖 7a-c) 。
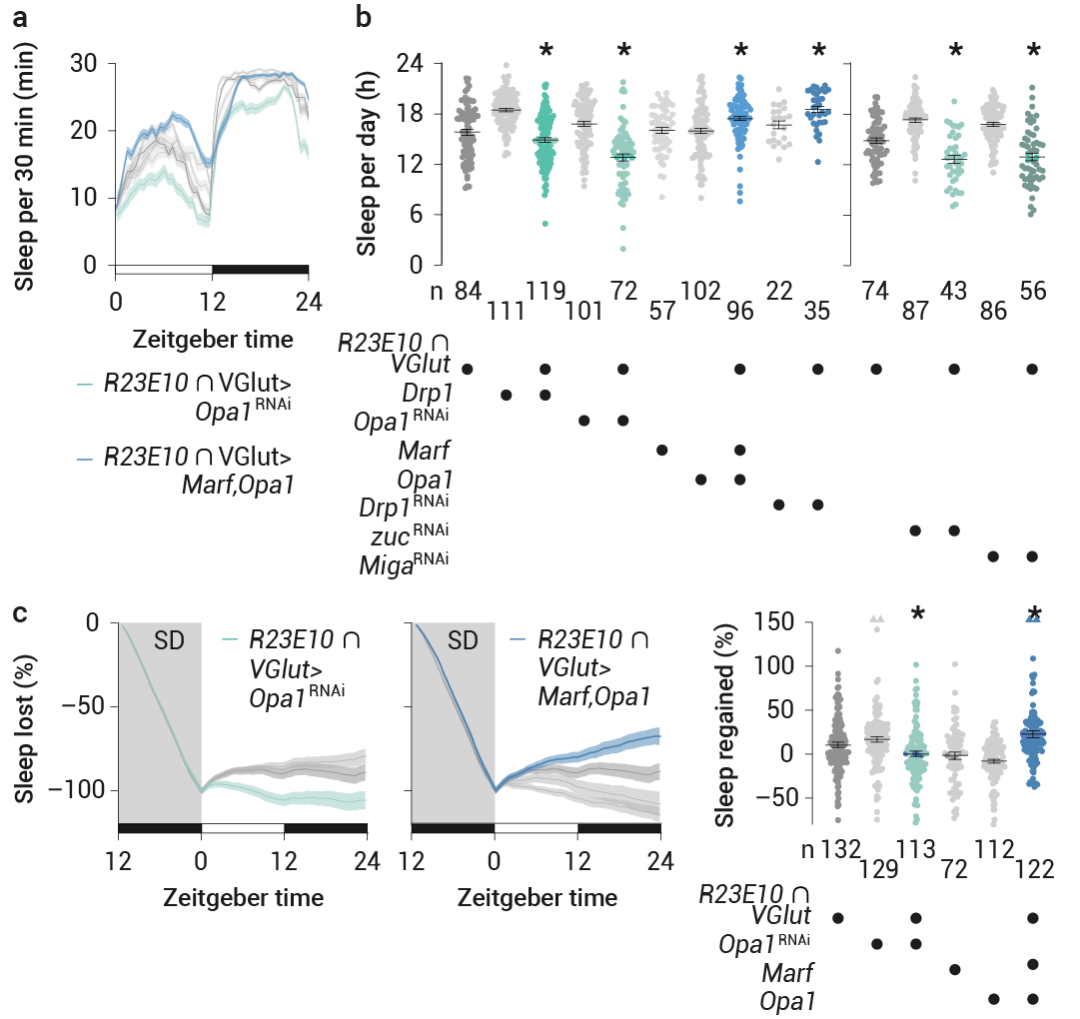 圖 7. 線粒體動力學改變睡眠
[5]
。
圖 7. 線粒體動力學改變睡眠
[5]
。
a-b. 表達 R23E10 ∩ VGlut-GAL4 驅動的裂變或融合蛋白的果蠅,或表達針對編碼這些蛋白質或調節磷脂酸水平的轉錄本的 RNAi 轉基因的果蠅?及其親本對照的睡眠概況 (a) 和每日睡眠 (b)。增加裂變 (綠色) 或融合 (藍色) 的操作會在相反的方向改變睡眠。c. 增加裂變(R23E10 ∩ VGlut-GAL4 > Opa1 RNAi,綠色)或融合 (R23E10 ∩ VGlut-GAL4 > Marf,Opa1,藍色) 的操作會改變時間進程 (左圖) 和睡眠剝奪后相反方向的反彈百分比 (右圖)。
最后,作者提到睡眠可能是為了滿足代謝需求而進化的。比如,線粒體融合后,AgRP 神經元的電輸出會增加,從而促進體重增加和脂肪沉積;這就像線粒體融合后 dFBN 的電輸出會增加以促進睡眠一樣。刪除 AgRP 神經元中的線粒體融合蛋白會損害食物攝入,就像干擾 dFBN 中的線粒體融合會損害睡眠誘導一樣。這些相似之處表明,睡眠壓力和饑餓感都源自線粒體,并且電子會流經各自反饋控制器的呼吸鏈,就像沙漏中的沙子一樣,決定何時需要恢復平衡。
Section.03
小結
總之,當你遲遲不睡時,你的線粒體率先“破大防”,碎片化線粒體增加,線粒體自噬增強,并頻繁“聯系”內質網,種種損傷只為喚起你的睡眠沖動,從而促使睡眠!當然,線粒體這些形態學變化在恢復睡眠后是可逆的,并且由于呼吸鏈中電子溢出的增加而減弱 (電子過剩誘導睡眠) 。此外,誘導或抑制 dFBN 中的線粒體裂變或融合,會以相反的方向改變睡眠和睡眠控制細胞的電特性:線粒體融合增加睡眠,而碎片化的線粒體減少睡眠。睡眠,像衰老一樣,可能是有氧代謝不可避免的結果。
| 產品推薦 |
| 是選擇性的發動蛋白相關蛋白 1 (Drp1) 抑制劑。Mdivi-1 是一種有效的線粒體分裂/線粒體自噬 (mitophagy) 抑制劑。 |
| 是一種線粒體融合 (mitochondrial fusion) 的小分子抑制劑。 |
| MitoTracker Deep Red/ MitoTracker Green (HY-D1783) 線粒體紅色/綠色熒光探針。 |
| 是一種廣泛用于檢測線粒體膜電位的理想熒光探針。正常線粒體內,JC-1 聚集在線粒體基質中形成聚合物,聚合物發出強烈的紅色熒光 (Ex=585 nm, Em=590 nm) ;在線粒體膜電位較低時,JC-1不能聚集在線粒體的基質中,產生綠色熒光 (Ex=510 nm, Em=527 nm) 。 |
| 是一種針對線粒體的熒光染料,能特異性靶向線粒體。TPE-PY 主要用于生物成像領域,特別是活細胞中線粒體的特異性染色研究,有助于觀察線粒體的位置、形態和數量。 |

[1] Ly S, Pack AI, Naidoo N. The neurobiological basis of sleep: Insights from Drosophila. Neurosci Biobehav Rev. 2018 Apr;87:67-86.
[2] 2017年度諾貝爾生理學或醫學獎, 生命科學, 2017 年 12 月, 第 29 卷 第 12 期, DOI: 10.13376/j.cbls/2017161
[3] Tainton-Heap LAL, et al. A Paradoxical Kind of Sleep in Drosophila melanogaster. Curr Biol. 2021 Feb 8;31(3):578-590.e6.
[4] Kato YS,et al. Interneurons of fan-shaped body promote arousal in Drosophila. PLoS One. 2022 Nov 21;17(11):e0277918.
[5] Kato YS, et al. Interneurons of fan-shaped body promote arousal in Drosophila. PLoS One. 2022 Nov 21;17(11):e0277918.


本站“ABIO生物試劑品牌網”圖片文字來自互聯網
如果有侵權請聯系微信: nanhu9181 處理,感謝~